




Первоначально этот переносчик присоединяет с внутренней стороны мембраны три иона Na +. Эти ионы изменяют конформацию активного центра АТФазы. После такой активации АТФаза способна гидролизовать одну молекулу АТФ, причем фосфат-ион фиксируется на поверхности переносчика с внутренней стороны мембраны.
Выделившаяся энергия расходуется на изменение конформации АТФазы, после чего три иона Na + и ион 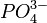 (фосфат) оказываются на внешней стороне мембраны. Здесь ионы Na + отщепляются, а замещается на два иона K +. Затем конформация переносчика изменяется на первоначальную, и ионы K + оказываются на внутренней стороне мембраны. Здесь ионы K + отщепляются, и переносчик вновь готов к работе.
(фосфат) оказываются на внешней стороне мембраны. Здесь ионы Na + отщепляются, а замещается на два иона K +. Затем конформация переносчика изменяется на первоначальную, и ионы K + оказываются на внутренней стороне мембраны. Здесь ионы K + отщепляются, и переносчик вновь готов к работе.
Более кратко действия АТФазы можно описать так:
- 1) Она изнутри клетки "забирает" три иона Na +,затем расщепляет молекулу АТФ и присоединяет к себе фосфат
- 2) "Выбрасывает" ионы Na + и присоединяет два иона K + из внешней среды.
- 3) Отсоединяет фосфат, два иона K + выбрасывает внутрь клетки
В итоге во внеклеточной среде создается высокая концентрация ионов Na +, а внутри клетки - высокая концентрация K +. Работа Na +, K + - АТФаза создает не только разность концентраций, но и разность зарядов (она работает как электрогенный насос). На внешней стороне мембраны создается положительный заряд, на внутренней - отрицательный.
12. Ядро. Строение и функции.
Клеточное ядро состоит из оболочки, ядерного сока, ядрышка и хроматина. Функциональная роль ядерной оболочки заключается в обособлении генетического материала (хромосом) эукариотической клетки от цитоплазмы с присущими ей многочисленными метаболическими реакциями, а также регуляции двусторонних взаимодействий ядра и цитоплазмы. Ядерная оболочка состоит из двух мембран, разделенных околоядерным (перинуклеарным) пространством. Последнее может сообщаться с канальцами цитоплазматической сети.
Ядерная оболочка пронизана порожу диаметром 80—90 нм. Область поры или поровый комплекс с диаметром около 120 нм имеет определенное строение, что указывает на сложный механизм регуляции ядерно-цитоплазматических перемещений веществ и структур. Количество пор зависит от функционального состояния клетки. Чем выше синтетическая активность в клетке, тем больше их число. Подсчитано, что у низших позвоночных животных в эритробластах, где интенсивно образуется и накапливается гемоглобин, на 1 мкм2 ядерной оболочки приходится около 30 пор. В зрелых эритроцитах названных животных, сохраняющих ядра, на 1 мк»г оболочки остается до пяти пор, т.е. в 6 раз меньше.
В области перового комплекса начинается так называемая плотная пластинка — белковый слой, подстилающий на всем протяжении внутреннюю мембрану ядерной оболочки. Эта структура выполняет прежде всего опорную функцию, так как при ее наличии форма ядра сохраняется даже в случае разрушения обеих мембран ядерной оболочки. Предполагают также, что закономерная связь с веществом плотной пластинки способствует упорядоченному расположению хромосом в интерфазном ядре.
Основу ядерного сока, или матрикса, составляют белки. Ядерный сок образует внутреннюю среду ядра, в связи с чем он играет важную роль в обеспечении нормального функционирования генетического материала. В составе ядерного сока присутствуют нитчатые, или фибриллярные, белки, с которыми связано выполнение опорной функции: в матриксе находятся также первичные продукты транскрипции генетической информации — гетероядерные РНК (гя-РНК), которые здесь же подвергаются процессингу, превращаясь в м-РНК (см. 3.4.3.2).
Ядрышко представляет собой структуру, в которой происходит образование и созревание рибосомальных РНК (рРНК). Гены рРНК занимают определенные участки (в зависимости от вида животного) одной или нескольких хромосом (у человека 13—15 и 21—22 пары) — ядрышковые организаторы, в области которых и образуются ядрышки. Такие участки в метафазных хромосомах выглядят как сужения и называются вторичными перетяжками. С помощью электронного микроскопа в ядрышке выявляют нитчатый и зернистый компоненты. Нитчатый (фибриллярный) компонент представлен комплексами белка и гигантских молекул РНК-предшественниц, из которых затем образуются более мелкие молекулы зрелых рРНК. В процессе созревания фибриллы преобразуются в рибонуклеопротеиновые зерна (гранулы), которыми представлен зернистый компонент.
Хроматиновые структуры в виде глыбок, рассеянных в нуклеоплазме, являются интерфазной формой существования хромосом клетки.
13. Цитоплазма. Органеллы общего значения и специальные, их строение и функции.
В цитоплазме различают основное вещество (матрикс, гиалоплазма), включения и органеллы. Основное вещество цитоплазмы заполняет пространство между плазмалеммой, ядерной оболочкой и другими внутриклеточными структурами. Обычный электронный микроскоп не выявляет в нем какой-либо внутренней организации. Белковый состав гиалоплазмы разнообразен. Важнейшие из белков представлены ферментами гаиколиза, обмена Сахаров, азотистых оснований, аминокислот и липидов. Ряд белков гиалоплазмы служит субъединицами, из которых происходит сборка таких структур, как микротрубочки.
Основное вещество цитоплазмы образует истинную внутреннюю среду клетки, которая объединяет все внутриклеточные структуры и обеспечивает взаимодействие их друг с другом. Выполнение матриксом объединяющей, а также каркасной функции может быть связано с выявляемой с помощью сверхмощного электронного микроскопа микротрабекулярной сети, образованной тонкими фибриллами толщиной 2—3 нм и пронизывающей всю цитоплазму. Через гиалоплазму осуществляется значительный объем внутриклеточных перемещений веществ и структур. Основное вещество цитоплазмы следует рассматривать так же, как сложную коллоидную систему, способную переходить из золеобразного (жидкого) состояния в гелеобразное. В процессе таких переходов совершается работа. О функциональном значении таких переходов.
Включениями называют относительно непостоянные компоненты цитоплазмы, которые служат запасными питательными веществами (жир, гликоген), продуктами, подлежащими выведению из клетки (гранулы секрета), балластными веществами (некоторые пигменты).
Органеллы — это постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции.
Выделяют органеллы общего значения и специальные. Последние в значительном количестве присутствуют в клетках, специализированных к выполнению определенной функции, но в незначительном количестве могут встречаться и в других типах клеток. К ним относят, например, микроворсинки всасывающей поверхности эпителиальной клетки кишечника, реснички эпителия трахеи и бронхов, синаптические пузырьки, транспортирующие вещества — переносчики нервного возбуждения с одной нервной клетки на другую или клетку рабочего органа, миофибриллы, от которых зависит сокращение мышцы. Детальное рассмотрение специальных органелл входит в задачу курса гистологии.
К органеллам общего значения относят элементы канальцевой и вакуолярной системы в виде шероховатой и гладкой цитоплазматической сети, пластинчатый комплекс, митохондрии, рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и микротрубочки, центриоли клеточного центра. В растительных клетках выделяют также хлоропласта, в которых происходит фотосинтез.
Канальцевая и вакуолярная системы образованы сообщающимися или отдельными трубчатыми или уплощенными (цистерна) полостями, ограниченными мембранами и распространяющимися по всей цитоплазме клетки. Нередко цистерны имеют пузыревидные расширения. В названной системе выделяют шероховатую и гладкую цитоплазматическую сети. Особенность строения шероховатой сети состоит в прикреплении к ее мембранам полисом. В силу этого она выполняет функцию синтеза определенной категории белков, преимущественно удаляемых из клетки, например секретируемых клетками желез. В области шероховатой сети происходит образование белков и липидов цитоплазматических мембран, а также их сборка. Плотно упакованные в слоистую структуру цистерны шероховатой сети являются участками наиболее активного белкового синтеза и называются эргастоплазмой.
Мембраны гладкой цитоплазматической сети лишены полисом. Функционально эта сеть связана с обменом углеводов, жиров и других веществ небелковой природы, например стероидных гормонов (в половых железах, корковом слое надпочечников). По канальцам и цистернам происходит перемещение веществ, в частности секретируемого железистой клеткой материала, от места синтеза в зону упаковки в гранулы. В участках печеночных клеток, богатых структурами гладкой сети, разрушаются и обезвреживаются вредные токсические вещества, некоторые лекарства (барбитураты). В пузырьках и канальцах гладкой сети поперечно-полосатой мускулатуры сохраняются (депонируются) ионы кальция, играющие важную роль в процессе сокращения.
Рибосома — это округлая рибонуклеопротеиновая частица диаметром 20—30 нм. Она состоит из малой и большой субъединиц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК). Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки бус. Такую структуру называют полисомой. Полисомы свободно располагаются в основном веществе цитоплазмы или прикреплены к мембранам шероховатой цитоплазматической сети. В обоих случаях они служат местом активного синтеза белка. Сравнение соотношения количества свободных и прикрепленных к мембранам полисом в эмбриональных недифференцированных и опухолевых клетках, с одной стороны, и в специализированных клетках взрослого организма — с другой, привело к заключению, что на полисомах гиалоплазмы образуются белки для собственных нужд (для «домашнего» пользования) данной клетки, тогда как на полисомах гранулярной сети синтезируются белки, выводимые из клетки и используемые на нужды организма (например, пищеварительные ферменты, белки грудного молока).
Пластинчатый комплекс Голъджи образован совокупностью диктиосом числом от нескольких десятков (обычно около 20) до нескольких сотен и даже тысяч на клетку.
Диктиосома представлена стопкой из 3—12 уплощенных дискообразных цистерн, от краев которых отшнуровываются пузырьки (везикулы). Ограниченные определенным участком (локальные) расширения цистерн дают более крупные пузырьки (вакуоли). В дифференцированных клетках позвоночных животных и человека диктиосомы обычно собраны в околоядерной зоне цитоплазмы. В пластинчатом комплексе образуются секреторные пузырьки или вакуоли, содержимое которых составляют белки и другие соединения, подлежащие выводу из клетки. При этом предшественник секрета (просекрет), поступающий.в диктиосому из зоны синтеза, подвергается в ней некоторым химическим преобразованиям. Он также обособляется (сегрегируется) в виде «порций», которые здесь же одеваются мембранной оболочкой. В пластинчатом комплексе образуются лизосомы. В диктиосомах синтезируются полисахариды, а также их комплексы с белками (гликопротеины) и жирами (гликолипиды), которые затем можно обнаружить в гликокаликсе клеточной оболочки.
Митохондрии — это структуры округлой или палочко-видной, нередко ветвящейся формы толщиной 0,5 мкм и длиной обычно до 5—10 мкм. В большинстве животных клеток количество митохондрий колеблется от 150 до 1500, однако в женских половых клетках их число достигает нескольких сотен тысяч. В сперматозоидах нередко присутствует одна гигантская митохондрия, спирально закрученная вокруг осевой части жгутика. Одна разветвленная митохондрия обнаружена в клетке такого паразита человека, как трипаносома.
Оболочка митохондрий состоит из двух мембран, различающихся по химическому составу, набору ферментов и функциям. Внутренняя мембрана образует впячивания листовидной (кристы) или трубчатой (тубулы) формы. Пространство, ограниченное внутренней мембраной, составляет матрикс органеллы. В нем с помощью электронного микроскопа обнаруживаются зерна диаметром 20—40 нм. Они накапливают ионы кальция и магния, а также полисахариды, например гликоген.
В матриксе размещен собственный аппарат биосинтеза белка органеллы. Он представлен 2—б копиями кольцевой и лишенной гистонов (как у прокариот) молекулы ДНК, рибосомами, набором транспортных РНК (тРНК), ферментами редупликации ДНК, транскрипции и трансляции наследственной информации. По основным свойствам: размерам и структуре рибосом, организации собственного наследственного материала — этот аппарат сходен с таковым у прокариот и отличается от аппарата биосинтеза белка цитоплазмы эукариотической клетки. Гены собственной ДНК кодируют нуклеотидные последовательности митохондриальных рРНК и тРНК, а также последовательности аминонокислот некоторых белков органеллы, главным образом ее внутренней мембраны. Аминокислотные последовательности (первичная структура) большинства белков митохондрий закодированы в ДНК клеточного ядра и образуются вне органеллы в цитоплазме.
Главная функция митохондрий состоит в ферментативном извлечении из определенных химических веществ энергии (путем их окисления) и накоплении энергии в биологически используемой форме (путем синтеза молекул аденозинтрифосфата — АТФ). В целом этот процесс называется окислительным (расформированием). В энергетической функции митохондрий активно участвуют компоненты матрикса и внутренняя мембрана. Именно с этой мембраной связаны цепь переноса электронов (окисление) и АТФ-синтетаза, катализирующая сопряженное с окислением фосфорилирование АДФ в АТФ. Среди побочных функций митохондрий можно назвать участие в синтезе стероидных гормонов и некоторых аминокислот (глутаминовая).
Лизосомы представляют собой пузырьки диаметром обычно 0,2—0,4 мкм, которые содержат набор ферментов кислых гидролаз, катализирующих при низких значениях рН гидролитическое (в водной среде) расщепление нуклеиновых кислот, белков, жиров, полисахаридов. Их оболочка образована одинарной мембраной, покрытой иногда снаружи волокнистым белковым слоем (на электронограммах «окаймленные» пузырьки). Функция лизосом — внутриклеточное переваривание оазличных химических соединений и структур.
Первичными лизосомами (диаметр 100 нм) называют неактивные органеллы, вторичными — органеллы, в которых происходит процесс переваривания. Вторичные лизосомы образуются из первичных. Они подразделяются на гетеролизосомы (фаголизосомы) и аутолизосомы (цитолизосомы). В первых переваривается материал, поступающий в клетку извне путем пиноцитоза и фагоцитоза, во вторых разрушаются собственные структуры клетки, завершившие свою функцию. Вторичные лизосомы, в которых процесс переваривания завершен, называют остаточными тельцами (телолизосомы). В них отсутствуют гидролазы и содержится непереваренный материал.
Микротельца составляют сборную группу органелл. Это ограниченные одной мембраной пузырьки диаметром 0,1—1,5 мкм с мелкозернистым матриксом и нередко кристаллоидными или аморфными белковыми включениями. К этой группе относят, в частности, пероксисомы. Они содержат ферменты оксидазы, катализирующие образование пероксида водорода, который, будучи токсичным, разрушается затем под действием фермента пероксидазы. Эти реакции включены в различные метаболические циклы, например в обмен мочевой кислоты в клетках печени и почек. В печеночной клетке число пероксисом достигает 70—100.
К органеллам общего значения относят также некоторые постоянные структуры цитоплазмы, лишенные мембран. Микротрубочки — трубчатые образования различной длины с внешним диаметром 24 нм, шириной просвета 15 нм и толщиной стенки около 5 нм. Встречаются в свободном состоянии в цитоплазме клеток или как структурные элементы жгутиков, ресничек, митотического веретена, центриолей. Свободные микротрубочки и микротрубочки ресничек, жгутиков и центриолей имеют разную устойчивость к разрушающим воздействиям, например химическим (колхицин). Микротрубочки строятся из стереотипных субьединиц белковой природы путем их полимеризации. В живой клетке процессы полимеризации протекают одновременно с процессами деполимеризации. Соотношением этих процессов определяется количество микротрубочек. В свободном состоянии микротрубочки выполняют опорную функцию, определяя форму клеток, а также являются факторами направленного перемещения внутриклеточных компонентов.
Микрофиламентами называют длинные, тонкие образования, иногда образующие пучки и обнаруживаемые по всей цитоплазме. Существует несколько разных типов микрофиламентов. Актиновые микрофиламенты благодаря присутствию в них сократимых белков (актин) рассматривают в качестве структур, обеспечивающих клеточные формы движения, например амебоидные. Им приписывают также каркасную роль и участие в организации внутриклеточных перемещений органелл и участков гиалоплазмы.
По периферии клеток под плазмалеммой, а также в околоядерной зоне обнаруживаются пучки микрофиламентов толщиной 10 нм — промежуточные филстенты. В эпителиальных, нервных, глиальных, мышечных клетках, фибробластах они построены из разных белков. Промежуточные филаменты выполняют, по-видимому, механическую, каркасную функцию.
Актиновые микрофибриллы и промежуточные филаменты, как и микротрубочки, построены из субъединиц. В силу этого их количество зависит от соотношения процессов полимеризации и деполимеризации.
Для животных клеток, части клеток растений, грибов и водорослей характерен клеточный центр, в состав которого входят центриоли. Центриолъ (под электронным микроскопом) имеет вид «полого» цилиндра диаметром около 150 нм и длиной 300—500 нм. Ее стенка образована 27 микротрубочками, сгруппированными в 9 триплетов. В функцию центриолей входит образование нитей митотического веретена, которые также образованы микротрубочками. Центриоли поляризуют процесс деления клетки, обеспечивая расхождение сестринских хроматид (хромосом) в анафазе митоза.
Митохондриальные болезни.
Начиная с конца 80-х годов XX века получены убедительные доказательства связи некоторых видов наследственной патологии у человека с мутациями митохондриальной ДНК (см. гл. 4.1) В зависимости от типа мутаций митохондриальные болезни разделяют на 4 группы:
а) болезни, вызванные точковыми мутациями, приводящими к замене консервативных аминокислот в собственных белках митохондрий. К ним относятся пигментный ретинит и нейроофтальмопатия Лебера, при которой наступает двусторонняя потеря зрения. Выраженность клинических признаков у больных этими заболеваниями коррелирует с количеством мутантной мтДНК, которое у разных больных может варьировать от 5 до 100% всей мтДНК;
б) болезни, вызванные мутациями в генах т-РНК, приводящими к многочисленным дегенеративным заболеваниям с различной степенью тяжести клинических проявлений, коррелирующей с количеством мутантной мтДНК;
в) болезни вызванные делениями и дупликациями участков митохондриалъных генов. У человека описано тяжелое заболевание молодого и среднего возраста — отсроченная кардиопатия, при которой обнаружены делеции мтДНК кардиоцитов. Заболевание носит семейный характер. В ряде случаев предполагается Х-сцепленное наследование, что позволяет думать о существовании ядерного гена, мутация которого вызывает делению до 50% мтДНК кардиоцитов;
г) болезни, вызванные снижением числа копий мтДНК, что является следствием определенных мутаций. К данной группе относятся летальная инфантильная дыхательная недостаточность и синдром молочнокислого ацидоза, при которых число копий мтДНК снижается до 1—2% от нормы. Снижение содержания мтДНК в клетках различных органов приводит к развитию миопатий, нефропатий, печеночной недостаточности и т.д. вследствие ослабления синтеза белков, кодируемых мтДНК.
Изменения в ДНК митохондрий сопровождаются нарушением их функций, связанных с клеточным дыханием. Это определяет характер и степень тяжести клинических проявлений митохондриалъных болезней.
Выдвинута также гипотеза о том, что накопление спонтанно возникающих мутаций мтДНК является звеном механизмов старения и развития дегенеративных процессов у человека.
Пероксисомные болезни — группа заболеваний, обусловленных нарушением структуры и функции пероксисом, - внутриклеточных органелл, присутствующих в каждой клетке организма за исключением зрелых эритроцитов. Около 50 ферментов пероксисом обеспечивают Р-окисление очень длинноцепочечных жирных кислот (ОДЦЖК), дикарбоновых кислот, пипеколиновой кислоты, простагландинов. В пероксисомах происходят начальные этапы биосинтеза плазмалогенов, входящих в структуру миелина и составляющих от 5% до 20% фосфолипидов клеточных мембран. Одна из важнейших функций пероксисом — зашита клетки от образующегося в ней атомарного кислорода с помощью химических превращений, в которых участвуют пероксисомные каталазы.
Возникновение пероксисомных болезней обусловлено нарушением сложных ферментативных реакций внутри органелл, процессов транспорта белков через мембраны органелл и функции их рецепторов. Белки, вовлеченные в биогенез пероксисом, называют пероксинами. В настоящее время определены функции более 20 пероксинов, для 9 из которых установлена хромосомная локализация кодирующих генов, обозначаемых РЕХ. В большинстве случаев патогенетические механизмы пероксисомных болезней связаны с нарушением четырех основных биохимических процессов — синтеза плазмалогенов, окисления ОДЦЖК и фитановой кислоты и деградации пипеколиновой кислоты. Особенностью пероксисомных болезней является возникновение нескольких заболеваний при мутациях в одном и том же гене, или появление одного клинического фенотипа, при наличии мутаций в разных генах пероксинов. Среди наиболее распространенных пероксисомных болезней гепато-церебро-ренальный синдром Цельвегера, ризомелическаяточечная хондродиетрофия, синдром Рефсума и адренолейкодистрофия. Частота пероксисомных болезней составляет): 25000-1:50000 новорожденных. На основании двух важных критериев — количества пероксисом в печени и степени нарушения их функций - эти болезни принято делить на три основные группы. К первой группе относятся пероксисомные болезни, при которых количество пероксисом в печени значительно снижено, а биохимические процессы в них нарушены. Во вторую группу входят заболевания, характеризующиеся нарушением лишь нескольких биохимических процессов в пероксисомах и нормальным их количеством в печени. При заболеваниях третьей группы биологическая функция пероксисом полностью подавлена, но их количество в печени соответствует норме.
Клиническая картина пероксисомных заболеваний включает: 1) черспно-лицевые аномалии; 2) патологию органов зрения (нарушение пигментации сетчатки, побледнение дисков зрительных нервов); 3) неврологические симптомы (мышечная гипотония, судороги, задержка раннего психомоторного развития); 4) гепатомегалию; 5) аномалии скелета (ризомелический тип укорочения конечностей).
Для демонстрации особенностей проявления пероксисомных болезней приводим клинико-генетические характеристики церебро-гепато-ренального синдрома Цельвегера, относящегося к первой группе.
Синдром Цельвегера.
Синдром Цельвегера объединяет группу генетически гетерогенных состояний. К клиническим проявлениям синдрома Цельвегера могут приводить мутации в генах пероксинов 1,2,3,5,6 и 12. Все варианты СЦ наследуются по аутосомно-рецессивному типу.
Первые симптомы отмечаются с рождения. Для больных характерна внутриутробная гипотрофия (вес при рождении не превышает 2500 г), дисморфизм в строении лица и черепа — увеличение размеров лба, монголоидный разрез глаз, периорбитальная полнота тканей, короткий вздернутый нос, микрогнатия. Среди наиболее типичных признаков: резкая мышечная гипотония, доходящая до атонии, и поликистоз почек. У всех больных отмечаются полиморфные пороки развития головного мозга. Часто диагностируется полимикрогирия, лизэнцефалия, агенезия мозолистого тела, очаги демиелинизации в белом веществе мозга, гидроцефалия. В ряде случаев выявляется патология глаз в виде врожденных катаракт и глауком, а также пороки сердца и наружных половых органов. Для заболевания характерна длительная желтуха и симптомы надпочечниковой недостаточности в первые месяцы жизни. У всех детей отмечается грубая задержка раннего психомоторного развития и снижение продолжительности жизни. Большинство больных погибает в течение первого года.
Лизосомные болезни накопления – это тяжелые наследственные заболевания обмена веществ, связанные с отсутствием лизосомальных ферментов. Недостаток этих ферментов приводит к тому, что макромолекулы (сложные комплексы белков, липидов и углеводов) не расщипляются и накапливаются в лизосомах. В результате сначала нарушается работа, клетки, затем тканей, а затем всего организма. Частота заболеваний этой группой генетических болезней составляет 1:5000 новорожденных. В зависимости от самого субстрата и группы поврежденных ферментов различают: сфинголипидозы (ганглиозидоз, болезнь Крабе, болезнь Гоше, метахроматическая лейкодистрофия,, болезнь Фарбера, болезнь Фабри,, болезнь Шиндлера, болезнь Нимана-Пика); муколипидозы и гликопротеинозы (цероидный липофусциноз, болезнь Вольмана, муколипидоз маннозидоз); мукополисахаридозы (синдром Гурлера, синдром Хантера, синдром Шейе, синдром Сан-Филиппо, синдром Морото -_Лами, синдром Моркио, синдром Слая).
Диагноз лизосомного заболевания можно заподозрить на основе внешних признаков: скелетные аномалии, грубые черты лица, а также умственной отсталости, поражений внутренних органов и систем. Манифестация этих симптомов может произойти как в период новорожденности, так и в уже зрелом возрасте.
Одно из самых известных лизосомных заболеваний – болезнь Гоше. В основе лежит незаменимого фермента бета-глюкоцереброзидазы, в результате чего мембранный жир накапливается в клетках Гоша с нарушением функций внутренних органов.
Если раньше диагноз «Болезнь Гоше» считался практически приговором, то сейчас при применении заместительной ферментотерапии имиглюцеразой у больных появилась возможность достигнуть нормальной жизни. При регулярном приеме препарата размеры печени и селезенки уменьшаются практически до нормального состояния, гемограмма нормализуется, изчезают боли в костях.
15. Дезоксирибонуклеи́новая кислота́ (ДНК) — макромолекула, обеспечивающая хранение, передачу из поколения в поколение и реализациюгенетической программы развития и функционирования живых организмов. Основная роль ДНК в клетках — долговременное хранение информации о структуре РНК и белков.
В молекуле ДНК присутствуют нуклеотиды четырех типов: дезоксиаденозин монофосфат (dAMP), дезоксигуанозинмонофосфат (dGMP), дезокситимидинмонофосфат(dТМР),дезоксицитадинмонофосфат(с!СМР). Номенклатура азотистых оснований, нуклеозидов и мононуклеотидов молекулы ДНК представлена в таблице.
ДНК имеет форму спирали, в которой основания разных цепей связаны между собой водородными связями. Цепи ДНК способны разделяться с помощью специальных ферментов и служить матрицами при синтезе дочерних молекул. Важнейшее свойство ДНК — комплементарность ее цепей. Это означает, что против аденина в одной из цепей всегда стоит тимин в другой цепи, гуанин всегда соединен с цитозином. Комплементарные пары аденин и тимин соединены двумя водородными связями, а гуанин с цитозином тремя водородными связями.
Помимо водородных связей между основаниями разных цепей стабильность двойной спирали ДНК обеспечивают гликозидные связи между азотистыми основаниями и остатками дезоксирибозы, а также фосфодиэфирные связи между двумя соседними остатками дезоксирибозы.
ДНК может существовать в виде нескольких форм, различающихся числом пар оснований на виток, утлом вращения между соседними парами оснований, расстоянием между парами оснований и диаметром спирали. В условиях in vivo наиболее частой является праюсторонняя В-форма, в которой одна цепь повернута вокруг другой по часовой стрелке. Имеется также и левосторонняя Z-форма.
Какие же из перечисленных выше структурных и функциональных особенностей молекулы ДНК позволяют ей хранить и передавать наследственную информации от клетки к клетке, от поколения к поколению, обеспечивать новые комбинации признаков у потомства?
1. Стабильность. Она обеспечивается водородными, гликозидными и фосфодиэфирными связями, а также механизмом репарации спонтанных и индуцированных повреждений;
2. Способность к репликации. Благодаря этому механизму в соматических клетках сохраняется диплоидное число хромосом. Схематично псе перечисленные особенности ДНК как генетической молекулы изображены на рисунке.
3. Наличие генетического кода. Последовательность оснований в ДНК с помощью процессов транскрипции и трансляции преобразуется в последовательность аминокислот в полипептидной цепи;
4. Способность к генетической рекомбинации. Благодаря этому механизму образуются новые сочетания сцепленных генов.
Передача генетической информации в клетке основана на матричных процессах (репликации, транскрипции, трансляции). Синтез дочерней цепи (репликация) молекулы ДНК происходит по матрице одной из двух родительских цепей с образованием новой двухиепочечной молекулы ДНК. Синтез молекулы РНК совершается в процессе транскрипции ДНК по матрице одной из двух цепей ДНК. Такая матричная (информационная) РНК может рассматриваться как посредник между ДНК и белком. Далее при синтезе белков генетическая информация, закодированная в последовательности триплетов азотистых оснований (канонов), транслируется в аминокислотную последовательность полипептидных цепей. Остановимся кратко на каждом из этих процессов,
Репликация. Во время репликации происходит расхождение двух цепей ДНК, и каждая из них служит матрицей для синтеза дочерней цепи. Такой способ репликации называется полуконсервативным. При этом дезоксирибонуклеотиды встраиваются в дочернюю цепь согласно правилу комплементарности азотистых оснований (А — Т, G — С). Вновь образованная молекула состоит из одной родительской и одной дочерней цепи ДНК. Образование дочерних хромосом происходит на стадии синтеза (S) в интерфазе между митотическими делениями и перед первым делением мейоза, В анафазе удвоенные хромосомы расходятся по дочерним клеткам. Таким образом, без процесса репликации невозможно сохранение диплоидного числа хромосом в соматических клетках и образование гаплоидного набора хромосом в половых клетках после двух делений мейоза. Однако при делении клеток происходит не только сохранение числа хромосом, но и воспроизведение последовательности азотистых оснований в молекулах ДНК, основанное на комплементарностb пар оснований родительской и дочерней цепей ДНК.
Цепи отделяются друг от друга, и каждая служит матрицей для построения комплементарной цепи. В результате синтезируются две молекулы, у каждой из которых одна цепь старая и одна новая. Такой способ репликации ДНК называют полуконсервативным.

