




3.2.1. Фоторегуляторные системы: механизм действия.
Виды ответных реакций на освещение
Вся эволюция животного и растительного мира проходила под мощным влиянием солнечного освещения, поэтому всё живое должно было приспособиться к такому постоянно действующему фактору.
Одноклеточная зелёная водоросль хламидомонада имеет так называемый «красный глазок», который воспринимает изменения в интенсивности освещённости, и клетка либо перемещается туда, где интенсивность света оптимальна для фотосинтеза, либо остаётся на месте, если освещённость достаточна. Такая ответная реакция на свет называется фототаксисом.
Одноклеточная зелёная водоросль эвглена также имеет глазок и фоторецептор, который обнаруживает источник света и заставляет клетку плыть в направлении оптимальной освещённости, что также является проявлением фототаксиса. Движение эвглены к свету, перемещение хлоропластов к свету – примеры положительного фототаксиса; избегание света дождевыми червями, личинками мясных мух, мокрицами, тараканами — примеры отрицательного фототаксиса.
Тропизмом называется ростовое движение растений под воздействием внешнего фактора. Фототропизм — ростовое движение растений под воздействием света: побеги обладают положительным, а придаточные корни — отрицательным фототропизмом.
Фотонастия — «сонные движения» некоторых цветков и листьев, когда они открываются или закрываются в ответ на изменение освещённости.
Животные также обладают фоторецепторами, реагирующими на свет. Все рецепторы животных преобразуют энергию стимула в локальный нераспространяющийся электрический сигнал, который возбуждает нервные импульсы в сенсорном нейроне. В центральной нервной системе импульсы расшифровываются и используются для выработки нужной ответной реакции. Время проявление эффекта определяется временем задержки ответной реакции.
Особенностью фоторегуляторных систем является их высокая квантовая чувствительность, сочетающаяся с большой эффективностью биологического действия. Это обусловлено триггерным принципом действия систем фоторегуляции: свет служит только сигналом, запускающим сложную цепь последовательных биохимических превращений, приводящих в конечном итоге к определённому фотобиологическому эффекту.
Пигменты–сенсибилизаторы большинства фоторегуляторных процессов до сих пор не идентифицированы за исключением фитохрома, универсально распространённого среди растительных организмов. Фитохром — хромопротеин, в котором хромофором служит линейный тетрапиррол (или билитриен). От взаимодействия хромофора и белковой части пигмента зависит фотообратимость и наличие двух взаимопревращающихся форм. Одна из форм имеет длинноволновый максимум поглощения при 660 нм, другая — при 730 нм. При фотоконверсии фитохрома происходит цис–транс изомеризация пигмента:

При переносе энергии электронного возбуждения с триплетного уровня триптофановых оснований в белке на синглетный возбуждённый уровень билитриена протекают процессы индуктивно–резонансной миграции энергии. Дезактивация синглетного возбужденного состояния фитохрома преимущественно происходит по фотохимическому пути в фотореакции изомеризации. Образованная при этом форма Ф730 может взаимодействовать с регуляторными и сигнальными молекулами.
Изучение действия фитохрома и некоторых фоторецепторов фоторегуля-торных реакций позволяет обобщить схему работы фоторегуляторных систем. Молекула фотохрома (условно назовём его так для общности) образует комп-лекс с ферментом (фотохром–протеин). Поглощая квант света, молекула фото-хрома претерпевает фотохимическую реакцию (например, цис–транс–изомери-зации), приводящую к изменению пространственной конфигурации фотохрома, что отражается на характере взаимодействия фотохрома с ферментом. Таким образом, общим свойством фотохромов является их способность изменять активность фермента за счёт непосредственного пространственно–структурного взаимодействия с его активным или аллостерическим центром.
В фототропических реакциях основным рецептором является флавиновые хромофоры. Они сенсибилизируют фотоокисление ауксина — гормона роста растений. Фототропические реакции имеют высокий квантовый выход (примерно 0,7) и высокую квантовую чувствительность: заметные изгибы растений наблюдаются при дозах синего света около 10-3 Дж/м2.
С помощью фототаксиса микроорганизмы регулируют уровень своей освещённости путём перемещения в области оптимального освещения для максимальной скорости фотосинтеза при минимальном повреждающем действии света. Для фототаксиса необходимо перемещение клетки параллельно световым лучам. При этом для детектирования направления распространения света используется анизотропия поглощения света фоторецептором, вероятно флавиновым пигментом.
3.2.2. Зрение как реакция на действие электромагнитного излучения оптического диапазона
Зрительное восприятие электромагнитного излучения. Глаз человека реагирует на электромагнитные колебания с длиной волны l = 400...760 нм. Наибольшую чувствительность глаз имеет к длине волны l=555 нм для дневного зрения, что соответствует максимуму солнечного излучения, попавшего на земную поверхность. Для сумеречного зрения максимум чувствительности глаза приходится на l=510 нм. При сумеречном зрении плохо различаются цвета, и излучение воспринимается как голубовато-серое.
Говоря о зрительном восприятииэлектромагнитного излучения, необходимо постоянно помнить о том, что с ним связано само понятие видимый спектр, т.е. в абсолютную шкалу электромагнитных волн властно вторглась биология. Как мы знаем из курса общей физики, благодаря наличию видимого спектра все фотометрические величины, т.е. величины, характеризующие перенос энергии электромагнитными волнами, делятся на энергетические и светотехнические. У них даже размерности разные, поскольку для светотехнических, т.е. привязанных к видимому спектру, величин, введена самостоятельная единица — кандела (Кд) и связанные с ней люмены (лм) и люксы (лк).
Связь между энергетическими и световыми величинами устанавливается через спектральную световую эффективность (иначе называемую кривой видности), определяемую как производную:
 (3.1)
(3.1)
В формуле (3.1) кривая видности вводится на основании сравнения энергетического и светотехнического потоков Ф и Ф е. Поскольку все остальные энергетические величины, характеризующие перенос энергии электромагнитными волнами, могут быть сопоставлены аналогичным образом, такое введение К l(λ) не ограничивает общности. Смысл кривой видности в том, что одинаковые по значению монохроматические потоки излучения разных длин волн вызывают различное зрительное ощущение и воспринимаются как разные цвета. Для практических оценок обычно пользуются не абсолютной кривой К l(λ), а относительной V (l), определяемой как отношение

 (3.2)
(3.2)
где K max– значение К l(λ) в максимуме зрительного ощущения. Это значение смещается при возрастании освещенности сетчатки (или яркости наблюдаемых предметов) в сторону длинных волн. Однако это смещение носит ограниченный характер: если яркость наблюдаемых объектов не превосходит 103 Кд/м2, то длина волны l (K max ) с точностью до справедливости модели усредненного глаза составляет 510 нм (сумеречное зрение). Если же яркость Ł возрастает до 104... 105 Кд/м2, что соответствует предметам, диффузно отражающим прямой солнечный свет, то l (K max ) смещается до 555 нм, но дальше не сдвигается вплоть до яркостей, нарушающих нормальное зрительное ощущение. Это явление ограниченного смещения максимума спектральной чувствительности глаза при переходе от сумеречного (Ł £ 103 Кд/м2) к дневному (Ł ~ 104...105 Кд/м2) освещению носит название эффекта Пуркинье.
Эффект Пуркинье вполне удовлетворительно объясняется особенностями строения чувствительных элементов сетчатки (см. ниже). Для нас сейчас важно выражение, позволяющее связать введенные фотометрические величины с их спектральными плотностями. Без ограничения общности запишем соответствующее выражение для потока:
 (3.3)
(3.3)
где интеграл берется по всему видимому спектру, φ e( λ) – спектральная плотность потока, а aj– коэффициент пересчета световых величин в энергетические, составляющий для lmax = 555 нм (случай дневного освещения) 625 лм/Вт. Световая эффективность в этом случае равна:
 . (3.4)
. (3.4)

| Ян (Иоганн) Евангелист Пуркинье (1787-1869) Чешский врач и физиолог, профессор анатомии и физиологии в Пражском и Бреславльском университетах. Труды по физиологии зрения, эмбриологии. |
Границы видимого диапазона, установленные как обобщение огромного количества субъективных измерений, имеют глубокий биофизический смысл. В процессе длительной эволюции глаз высших позвоночных и человека практически идеально приспособился к восприятию дневного и сумеречного освещения. На видимый спектр приходится около 40% энергии солнечного излучения, хотя объективно это весьма узкий интервал (менее 1 октавы). Вблизи поверхности Земли солнечный спектр обрывается на 290 нм. Более короткие длины волн задерживаются слоем озона (см. выше). Тем самым восприятие длин волн короче 300 нм было бы нецелесообразно как с чисто физической точки зрения (их почти не пропускает атмосфера), так и с биологической (излучение с длинами волн короче 300 нм вызывает фотохимические реакции, нарушающие естественные метаболические процессы, причем тем более сильно, чем короче длина волны). Казалось бы, мы сталкиваемся с противоречием: сам факт регистрации света органом зрения есть цепь фотохимических реакций. Однако эти реакции обратимы, а от реакций, вызываемых более жестким излучением и носящих необратимый характер, необходима защита. Поэтому максимум чувствительности сетчатки глаза лежит вблизи границы видимого диапазона (380 нм), но поглощение в хрусталике смещает общий максимум кривой видности в зеленую область. Тем самым передние отделы глаза обеспечивают должную защиту от нежелательных фотохимических реакций, вызываемых УФ-излучением с длиной волны l<380 нм, но не препятствуют желательным. Медицинская практика свидетельствует о помутнении хрусталика при длительном облучении жестким УФ-излучением (защитные возможности хрусталика не беспредельны), а в случае удаления помутневшего хрусталика происходит заметный сдвиг кривой видности в коротковолновую область: пациент воспринимает все объекты как бы в голубом тумане (если только глаз продолжает функционировать).
Касательно длинноволновой границы видимого диапазона отметим, что таковая характеризуется на порядок меньшей величиной V (l), чем коротковолновая. Это не случайно. Высокая чувствительность глаза к ИК излучению была бы не только нецелесообразна, но вообще не дала бы возможности ничего видеть при солнечном освещении. Причина — в собственном тепловом излучении глаза. При сравнительно низкой температуре глаза (Т ~ 310 К) все его тепловое излучение приходится на ИК область. Оценим по закону Стефана-Больцмана (см.ниже) мощность, излучаемую внутрь глаза: 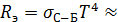
≈ 0,105 Вт/см2 (здесь обозначено: R э— интегральная плотность мощности, или энергетическая светимость глаза как источника теплового излучения, σС-Б — постоянная Стефана-Больцмана.) Общая внутренняя поверхность усредненного глаза составляет около 17 см2, поэтому полная излучаемая внутрь глаза мощность (лучистый поток)  1,7 Вт. Оценим теперь мощность прямого солнечного излучения, попадающего в глаз через зрачок. Солнечная постоянная, т.е. усредненный поток, падающий на единицу поверхности Земли перпендикулярно направлению на Солнце составляет около 0,139 Вт/см2. При радиусе зрачка r
1,7 Вт. Оценим теперь мощность прямого солнечного излучения, попадающего в глаз через зрачок. Солнечная постоянная, т.е. усредненный поток, падающий на единицу поверхности Земли перпендикулярно направлению на Солнце составляет около 0,139 Вт/см2. При радиусе зрачка r 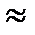 2 мм (оценка завышена) имеем Фсолн 0,139p r2 » 0,017 Bт. Это в 100 (!) раз меньше собственного теплового излучения глаза. Таким образом, если бы глаз в ИК области был бы столь же чувствителен, как и в видимой, то собственное тепловое излучение забило бы даже прямой солнечный свет. Интересно, что при всей простоте приведённой оценки она впервые была проделана относительно недавно С.И. Вавиловым. Это биологический аспект резкого спада чувствительности глаза в инфракрасной области.
2 мм (оценка завышена) имеем Фсолн 0,139p r2 » 0,017 Bт. Это в 100 (!) раз меньше собственного теплового излучения глаза. Таким образом, если бы глаз в ИК области был бы столь же чувствителен, как и в видимой, то собственное тепловое излучение забило бы даже прямой солнечный свет. Интересно, что при всей простоте приведённой оценки она впервые была проделана относительно недавно С.И. Вавиловым. Это биологический аспект резкого спада чувствительности глаза в инфракрасной области.
Другая причина — чисто физическая: процесс регистрации электромагнитного излучения органом зрения есть процесс фотохимический, т.е. цепочка реакций, вызываемых отдельными квантами. Если энергия кванта ниже некоторой пороговой величины, то инициировать эту цепь реакций он не в состоянии (аналогично красной границе фотоэффекта).

| Рис. 3.4. Схематическое строение глаза (обозначения по Гульстранду). |
Глаз окружен плотной непрозрачной оболочкой ww (склера). Через заднюю стенку склеры проходит глазной нерв е и кровеносные сосуды. Передняя часть склеры переходит в прозрачную роговицу h, более выпуклую, чем 
| Альвар Гульстранд (1862-1930) Шведский офтальмолог, лауреат Нобелевской премии по физиологии и медицине (1911) за цикл работ по оптике зрения. Профессор офтальмологии Упсальского университета (с 1894), создатель и руководитель кафедры физической и физиологической оптики Упсальского университета (с 1914 по 1927). Член Нобелевского комитета по физике с 1911 по 1929. Автор фундаментальных исследований по оптике органа зрения, разработчик средств коррекции дефектов зрения. Лауреат многих почетных премий и наград, обладатель ряда ученых степеней в разных странах Европы (Швеции, Германии, Ирландии). |
склера. Толщина склеры составляет от 0,4 до 1,1 мм, роговицы — 0,5 мм. Проведем оптическую ось глаза АВ. Двигаясь внутрь глаза по направлению от роговицы h, попадем сначала в переднюю глазную камеру a, затем в хрусталик
 и заднюю глазную камеру g
и заднюю глазную камеру g  . Эти части вместе с роговицей составляют оптическую систему глаза, дающую изображения всех предметов. Толщины передней глазной камеры и хрусталика примерно одинаковы (3,6 мм). Передняя глазная камера заполнена жидкостью, задняя — студнеобразным веществом (стекловидное тело). Показатели преломления — как жидкости, так и стекловидного тела — близки к физраствору и составляют 1,336. Показатель преломления роговицы — 1,376, внутренняя поверхность склеры покрыта сосудистой оболочкой. Спереди она переходит в утолщенную часть st с кольцевидной ресничной мышцей uu и радужной оболочкой ii, окраска которой индивидуальна. Посредине радужки имеется зрачок Р, играющий роль апертурной диафрагмы. Изображение этого фактического отверстия, даваемое расположенными перед ним роговицей и передней камерой, играет роль входного зрачка, а изображение, даваемое хрусталиком и стекловидным телом — выходного зрачка оптической системы. Для практических оценок можно считать, что оба эти изображения совпадают с действительным зрачком. В зависимости от величины яркости наблюдаемых объектов диаметр зрачка может меняться за счет усилий мышцы uu в пределах от 2 до 8 мм и тем самым регулировать величину светового потока, попадающего в глаз.
. Эти части вместе с роговицей составляют оптическую систему глаза, дающую изображения всех предметов. Толщины передней глазной камеры и хрусталика примерно одинаковы (3,6 мм). Передняя глазная камера заполнена жидкостью, задняя — студнеобразным веществом (стекловидное тело). Показатели преломления — как жидкости, так и стекловидного тела — близки к физраствору и составляют 1,336. Показатель преломления роговицы — 1,376, внутренняя поверхность склеры покрыта сосудистой оболочкой. Спереди она переходит в утолщенную часть st с кольцевидной ресничной мышцей uu и радужной оболочкой ii, окраска которой индивидуальна. Посредине радужки имеется зрачок Р, играющий роль апертурной диафрагмы. Изображение этого фактического отверстия, даваемое расположенными перед ним роговицей и передней камерой, играет роль входного зрачка, а изображение, даваемое хрусталиком и стекловидным телом — выходного зрачка оптической системы. Для практических оценок можно считать, что оба эти изображения совпадают с действительным зрачком. В зависимости от величины яркости наблюдаемых объектов диаметр зрачка может меняться за счет усилий мышцы uu в пределах от 2 до 8 мм и тем самым регулировать величину светового потока, попадающего в глаз.
Сетчатка глаза имеет весьма сложное строение, схематически показанное на рис. 3.5. Она состоит из нескольких слоев нервных клеток различного типа и назначения. Все они решают задачу преобразования оптического изображения в нейральное, т.е. специальным образом закодированный набор нервных импульсов. В последнем слое находятся рецепторные клетки — палочки и колбочки. На выходе палочек и колбочек получаются электрические импульсы, передающиеся в мозг. Длина палочек составляет 0,05…0,08 мм, колбочек — 0,035 мм. Размер палочек 50×30 мкм. Общее число палочек в сетчатке около 130 млн, колбочек — 7 млн (тем самым, соотношение между числом палочек и колбочек составляет примерно 20:1). Периферия сетчатки заполнена почти исключительно палочками. По мере приближения к желтому пятну концентрация колбочек возрастает, и в пределах центральной ямки (в круге диаметром около 0,2 мм) палочек нет, там находятся только колбочки. Жёлтым пятно называется потому, что при изучении глаза через щелевую лампу это пятно на белом фоне сетчатки визуально воспринимается как жёлтое. В месте входа глазного нерва нет ни палочек, ни колбочек. Это место называется слепым пятном.
Поле зрения, соответствующее желтому пятну, невелико (6° по горизонтали, 4° по вертикали). Центральная ямка составляет около 1° по горизонтали и вертикали. Глаз обладает способностью быстро поворачиваться в глазной впадине, сканируя всю видимую поверхность большого предмета. Поэтому поле ясного зрения соответствует примерно 150° по горизонтали и 120° по вертикали. Вместе с тем подвижность глаза позволяет концентрировать внимание на наиболее важных деталях изображения.
Функции палочек и колбочек. Палочки обладают бόльшей по сравнению с колбочками светочувствительностью и реагируют на слабое освещение. Палочка способна генерировать зрительный сигнал в ответ на поглощение даже одного кванта света. При освещенности менее 10-2 лк зрительное ощущение вызывается только палочками. Они содержат только один зрительный пигмент (родопсин) и в силу этого не способны различать цвета. Поэтому сумеречное зрение обусловлено преимущественно палочками.

На наружном сегменте палочки световая энергия преобразуется в рецепторный потенциал (рис. 3.6). Наружный сегмент полностью состоит из мембран. В палочках число мембранных дисков достигает 600...1000. От внутреннего сегмента через синаптический участок ряд структур ведёт к зрительному нерву.
Колбочки содержат по крайней мере три зрительных пигмента, что обеспечивает восприятие цвета. Каждый цвет представляет собой комбинацию сигналов, посылаемых каждым из пигментов в ответ на раздражение излучением с заданной длиной волны, т.е. фотоном заданной энергии. Ощущение данного цвета есть, тем самым, декодировка импульсного сигнала, включающего три импульса с заданным соотношением амплитуд. Колбочки работают главным образом при дневном освещении. Цветное зрение начинается с освещенностей порядка 1 лк. При освещённости более 102 лк зрение осуществляется почти исключительно колбочками. Зрительные пигменты колбочек характеризуемых различными кривыми поглощения (рис. 3.7).
Палочковое зрение отличается меньшей остротой, так как палочки расположены менее плотно, равномерно по всей сетчатке, кроме центральной ямки, где сосредоточены в основном колбочки, дающие остроту зрения своей плотной упаковкой. Детали предмета различаются наиболее отчетливо, когда изображение получается на желтом пятне и, в особенности, на центральной ямке (фовеальное зрение). Периферическое зрение, напротив, имеет место при расположении изображения вдали от центральной ямки. Две близкие светящиеся точки глаз воспринимает раздельно только при условии, что угловое расстояние dJ между этими точками не меньше определенного предела, называемого предельным разрешаемым угловым расстоянием, а обратная ему величина называется разрешающей способностью, или просто остротой зрения. Нормальная острота зрения соответствует предельному угловому расстоянию порядка одной минуты. Острота зрения максимальна, если изображение попадает на центральную ямку.

| Рис. 3.7. Кривые поглощения зрительных пигментов для приматов (имеют большое сходство с кривыми поглощения зрительных пигментов человека). Максимумы кривых приходятся на 447 нм (сине-фиолетовый), 540 нм (зелёный), 577 нм (жёлтый). |
В самом деле, для разрешения необходимо, чтобы изображения соседних точек приходились на разные колбочки, поскольку мозг взаимно однозначно реагирует на раздражение каждой колбочки. Палочковое же зрение устроено так, что с одним волокном зрительного нерва соединено много палочек, поэтому оно увеличивает чувствительность глаза в условиях слабой освещенности, за что приходится платить уменьшением разрешения и невозможностью различать цвета. Поэтому разрешение максимально в центральной ямке, где можно пренебречь наличием палочек. Угловое разрешение dJ равно углу, под которым видно из задней узловой точки глаза (точка ƒ на рис 3.4) среднее расстояние d х между двумя соседними колбочками на центральной ямке, т.е. dJ
d х/f, где f — переднее фокусное расстояние глаза. Для глаза d х 0,0025 мм (2,5 мкм), f 15 мм (в среднем по Гульстранду), отсюда dJ 30".
Учитывая изложенное, можно дать объяснение эффекту Пуркинье. При малых яркостях свет воспринимается только палочками, при возрастании яркости все больше включаются колбочки. Одновременно при росте яркости Ł уменьшается зрачок, и изображение все больше концентрируется в области желтого пятна и центральной ямки. При Ł > 105 кд/м2 изображение сосредоточено в области центральной ямки, и в дальнейшем какие-либо изменения в восприятии отсутствуют (по крайней мере до тех пор, пока не нарушится нормальный ход реакций, связанных с работой колбочек).
Зрительная адаптация.Светочувствительностью глаза называют величину, обратную пороговой яркости, т.е. минимальной яркости, вызывающей зрительное ощущение в данных условиях наблюдения. Она изменяется в широких пределах (от 10-7 до 105 кд/м2, т.е. от 10-17 до 10-5 Вт) благодаря зрительной адаптации — способности глаза приспосабливаться к различному уровню яркости.
При понижении яркости происходит темновая, при повышении — световая адаптация. Темновая адаптация происходит значительно медленнее, чем световая. Механизм адаптации выяснен только частично.
Адаптация осуществляется следующими способами:
1) изменением диаметра зрачка в пределах от 2 до 8 мм, что изменяет световой поток в 16 раз, и за счет изменения площади изображения на сетчатке;
2) изменением количества светочувствительных элементов, участвующих в образовании нервного импульса:
Þ объединением светочувствительных элементов в один канал;
3) экранированием колбочек и палочек тёмным пигментом;
4) изменением концентрации светочувствительных пигментов, которые под действием света частично разлагаются, а затем восстанавливаются. При этом происходит изменение чувствительности самих рецепторов (главным образом палочек).
Установлено, что чувствительность палочек обусловлена в основном наличием светочувствительного пигмента родопсина, разлагающегося на свету и вновь восстанавливающегося в темноте. Чувствительность палочек, тем самым, меняется в зависимости от концентрации восстановленного родопсина. Спектральная кривая поглощения родопсина близка к сумеречной кривой видности. По всей вероятности, темновая адаптация (изменение чувствительности при падении яркости объекта от 103 кд/м2 до темноты примерно в 107 раз) главным образом обусловлена изменением концентрации родопсина в палочках.
Чувствительность колбочек меняется в значительно меньших пределах (примерно в 102 раз). Кроме того, все больше фактов свидетельствует о том, что изменение чувствительности рецепторов не объясняет полностью адаптацию глаза. Дело в том, что динамика изменения концентрации родопсина не совпадает с динамикой собственно адаптации. В частности, наибольшее изменение чувствительности глаза приходится на сравнительно малый начальный отрезок времени, когда изменение концентрации родопсина еще незначительно. Напротив, в последующий длительный период, когда концентрация родопсина меняется сильно, изменение чувствительности незначительно. Этот эффект не нашел пока удовлетворительного объяснения.
Нижний предел (абсолютный порог) чувствительности глаза при полной темновой адаптации составляет в среднем около 50...100 фотонов/с. Из них не более 10% поглощаются молекулами зрительного пигмента в палочках сетчатки, остальные отражаются от роговицы, поглощаются оптическими средам глаза или пигментным эпителием. Так что зрительная регистрация отдельных фотонов — это реальность.
Механизм фоторецепции палочек. Для колбочек, содержащих зрительный пигмент иодопсин (причем, возможно, не единственный), механизм фоторецепции изучен пока недостаточно. Поэтому остановимся более подробно на механизме фоторецепциипалочек.
Светочувствительный пигмент родопсин (устаревшее название — зрительный пурпур), представляющий собой половину молекулы b-каротина, находится на наружной поверхности мембранных дисков. Мембранные диски представляют собой двухслойные мембраны. Электрические оси молекул родопсина в них ориентированы, вообще говоря, хаотически, но в силу строения мембранных дисков эта хаотичность не является абсолютной (см. ниже). Родопсин представляет собой сложную молекулу, образующуюся в результате обратимого связывания липопротеина скотопсина с молекулой каротиноида ретиналя. Диаметр молекулы родопсина около 4 нм, объем ~ 0,75×10-19 см3. Молекулярный вес около 40000.
Ретиналь поглощает свет. Он представляет собой альдегидную форму витамина А и может в зависимости от освещения существовать в виде двух изомеров: 11- цисретиналь и полностью трансретиналь.
На сегодня принимается, что механизм фоторецепции включает 2 процесса: 1) превращение 11-цисретиналя в трансретиналь (фотоизомеризация) под действием кванта света; 2) расщепление родопсина через ряд промежуточных продуктов на ретиналь и скотопсин (выцветание).
Цикл цистрансизомеризации является обязательной частью любой известной фоторецепторной системы. Время жизни молекулы ретиналя в возбуждённом состоянии 10-9 с; время, нужное для поворота связи — от 10-11 до 10-12 с, что много меньше, чем в растворах. Это имеет место ввиду специфического белкового окружения ретиналя. Квантовый выход для реакции изомеризации 11-цис ® полностьютранс составляет 0,5-0,7.
За изомеризацией хромофора следует изменение структуры липопротеина — опсина. Конформационное изменение опсина приводит в конечном итоге к возникновению нервного импульса. При этом родопсин переходит с междисковой гидрофильной поверхности на внутреннюю гидрофобную фазу мембраны. В результате происходит увеличение проницаемости мембраны для ионов К+ и уменьшение для ионов Na+. Поглощение одного кванта света приводит к закрытию от 100 до 300 натриевых каналов. Если в состоянии покоя разность потенциалов между протоплазмой палочки и наружной средой –40 мВ, то освещение вызывает гиперполяризацию клеточной мембраны до –70...–80 мВ. Это приводит к возникновению потенциалов, вызывающих нервный импульс. Особенностью наружных сегментов палочек является то, что в темноте потенциал имеет натриевую природу, а под действием света — становится калиевой природы. Поэтому, в отличие от других известных клеток, на цитоплазматической мембране наружных сегментов палочек потенциал имеет знак «+» внутри и знак «–» снаружи. При этом процесс формирования сигнала в палочках под действием света является обратным процессу формирования нервного импульса, например, в нервных клетках.
После прекращения действия света родопсин ресинтезируется. Вначале полностью трансретиналь при участии фермента ретиналь-изомеразы превращается в 11-цисретиналь, а затем последний соединяется со скотопсином. Этот процесс лежит в основе темновой адаптации. В полной темноте требуется около 30 минут, чтобы все палочки адаптировались и глаз приобрел максимальную чувствительность.
Описанные процессы, дающие представление о сложных фотохимических превращениях с участием родопсина, не дают достаточного объяснения аномально высокой чувствительности глаза в полной темноте. Поэтому рассмотрим более подробно процесс регистрации света в смысле преобразования определенной порции энергии в электрический импульс, передающийся затем по нервному волокну в мозг.
Вообще молекула родопсина имеет сложную разветвленную структуру, но внутри мембранного диска ввиду плотной упаковки ее можно представлять себе в виде шарика. Полагая, что прохождение света сквозь слой молекул родопсина описывается законом Бугера-Ламберта-Бера  (см. Л2), можно оценить натуральный показатель поглощения: aп= n σ, где σ = 10-16 см2, n = 1019 см-3 (считая, что фотон поглощается одним из атомов в молекуле родопсина с поперечником 10-8 см и площадью ~ 10-16 см2; концентрация 1019 см-3 соответствует плотной упаковке шариков с диаметром ~ 5·10-7 см). Тем самым имеем натуральный показатель поглощения aп = n σ = 103 см-1, и поглощение происходит в слое толщиной около 10 мкм. Заметим также, что при малой толщине дисков дипольные моменты молекул родопсина всегда оказываются перпендикулярны направлению распространения света, что наиболее благоприятно для максимальной эффективности взаимодействия падающего излучения с какой-то из них, поскольку электрический вектор в волне лежит в одной плоскости с дипольным моментом молекулы. При этом, поскольку ориентация дипольных моментов молекул родопсина случайна, глаз должен примерно вдвое слабее реагировать на плоско-поляризованный свет, чем на естественно поляризованный. Количество дисков во внешнем сегменте палочки велико (размер активной зоны составляет около 40 мкм, так что поглощение фотона гарантировано). Если принять толщину диска 0,025 мкм, то при плотной упаковке дисков имеем примерно 1,6. 103 дисков с толщиной мембран 30 нм, дающих протяженность пути фотона внутри палочки около 0,5 мкм. В таком слое должно поглотиться примерно 99% интенсивности падающего света. Итак, фотон с вероятностью, близкой к 1, поглощается фоточувствительным ферментом палочки. Но отсюда еще не следует, что нервное волокно почувствует соответствующий импульс. Ведь энергия одного фотона зеленого света h ν = 1,24/0,5 = 2,5 эВ, достаточная для возбуждения одного атома в молекуле родопсина с молекулярной массой М ~ 40000, сама по себе ничтожна для того, чтобы влиять на состояние молекулы в целом и тем более на мембранный потенциал диска (в одном диске около 106 молекул родопсина). Тем не менее, при поглощении фотона мембранный потенциал палочки меняется на 40 мВ (!), т.е. дело обстоит так, как если бы один поглощенный фотон вызывал появление 106...107 ионов. Здесь явно присутствует спусковой механизм, работа которого была изучена и описана только в последнее время (рис. 3.8).
(см. Л2), можно оценить натуральный показатель поглощения: aп= n σ, где σ = 10-16 см2, n = 1019 см-3 (считая, что фотон поглощается одним из атомов в молекуле родопсина с поперечником 10-8 см и площадью ~ 10-16 см2; концентрация 1019 см-3 соответствует плотной упаковке шариков с диаметром ~ 5·10-7 см). Тем самым имеем натуральный показатель поглощения aп = n σ = 103 см-1, и поглощение происходит в слое толщиной около 10 мкм. Заметим также, что при малой толщине дисков дипольные моменты молекул родопсина всегда оказываются перпендикулярны направлению распространения света, что наиболее благоприятно для максимальной эффективности взаимодействия падающего излучения с какой-то из них, поскольку электрический вектор в волне лежит в одной плоскости с дипольным моментом молекулы. При этом, поскольку ориентация дипольных моментов молекул родопсина случайна, глаз должен примерно вдвое слабее реагировать на плоско-поляризованный свет, чем на естественно поляризованный. Количество дисков во внешнем сегменте палочки велико (размер активной зоны составляет около 40 мкм, так что поглощение фотона гарантировано). Если принять толщину диска 0,025 мкм, то при плотной упаковке дисков имеем примерно 1,6. 103 дисков с толщиной мембран 30 нм, дающих протяженность пути фотона внутри палочки около 0,5 мкм. В таком слое должно поглотиться примерно 99% интенсивности падающего света. Итак, фотон с вероятностью, близкой к 1, поглощается фоточувствительным ферментом палочки. Но отсюда еще не следует, что нервное волокно почувствует соответствующий импульс. Ведь энергия одного фотона зеленого света h ν = 1,24/0,5 = 2,5 эВ, достаточная для возбуждения одного атома в молекуле родопсина с молекулярной массой М ~ 40000, сама по себе ничтожна для того, чтобы влиять на состояние молекулы в целом и тем более на мембранный потенциал диска (в одном диске около 106 молекул родопсина). Тем не менее, при поглощении фотона мембранный потенциал палочки меняется на 40 мВ (!), т.е. дело обстоит так, как если бы один поглощенный фотон вызывал появление 106...107 ионов. Здесь явно присутствует спусковой механизм, работа которого была изучена и описана только в последнее время (рис. 3.8).
Описанный выше механизм закрытия натриевых и открытия калиевых каналов мембраны требует более подробного описания для выявления спусковых явлений. Именно, в мембране внешнего сегмента палочки имеются каналы, по которым ионы Na+ могут переноситься внутрь. В темноте через каналы внешнего сегмента идет ионный ток, и мембранный потенциал снижается (в стационарном темновом состоянии он составляет –40 мВ) (см. рис.3.8а). На свету возбуждение атома в молекуле родопсина запускает реакцию выделения медиатора (ряд процессов, важнейшую роль в которых играет белок трансдуцин: родопсин ® трансдуцин ® фосфодиэстераза ® гидролиз циклогуанинмонофосфата (цГМФ)). Последняя фаза этой цепочки реакций — гидролиз цГМФ фосфодиэстеразой — и приводит к закрытию натриевых каналов внешней мембраны. В результате ток ионов Na+ падает, и потенциал внутренней мембраны возрастает до –80 мВ (см. рис.3.8б). Характерное время реакции составляет около 1 мс.

| а) б) Рис. 3.8. Схема работы спускового механизма палочки: а) в темноте идет ионный ток, цепь замкнута; б) на свету цепь размыкается, ионный ток практически прекращается, потенциал возрастает |
Итак, поглощение фотона молекулой родопсина само по себе всего лишь активирует один атом в молекуле. Но активированная вследствие этого молекула родопсина запускает лавинообразный каскад процессов, благодаря которым происходит гиперполяризация внутреннего сегмента палочки и выработка благодаря этому миллисекундного нервного импульса. Этот механизм, связанный с закрытием мембранных Na+–каналов во внешнем сегменте палочки, исследован впервые Е.Е. Фесенко с сотрудниками в 1985-86 г.г. и подтвержден экспериментами с использованием электронной микроскопии, электронного парамагнитного резонанса, рентгенографии, эффекта Мёссбауэра (гамма-резонансной спектроскопии). Заметим, что гиперполяризованная палочка, характеризующаяся избытком внутриклеточного медиатора, приходит в нечувствительное состояние, поскольку родопсин в ходе каскадных превращений, вызывающих гиперполяризацию, распадается на фрагменты. Тем самым, чем больше света поглощено, тем больше молекул в диске изначально активизировано и тем гарантированнее остановка реакций в итоге распада активизированных молекул. Но как только процесс распада родопсина остановится, выделение внутриклеточного медиатора тоже прекратится. Это вызовет открытие каналов и возобновление транспорта Na+. Как только транспорт возобновится, причина, вызывающая распад родопсина, ликвидируется. При этом количество молекул родопсина в одном диске как раз соответствует количеству распавшихся молекул на одну активизированную. Палочка теряет полностью чувствительность при поглощении примерно 103 фотонов (количество дисков). Тем самым динамический диапазон (соотношение между максимальной и минимальной регистрируемыми интенсивностями) для палочек составляет 103¸104 при общем количестве палочек около 130 млн, хотя общее количество нервных волокон, связанных с палочками, порядка 10 млн. (примерно такое же, как и для колбочек), что приводит к дополнительному увеличению амплитуды нервного импульса, передаваемого в мозг.
Заметим, что процесс гиперполяризации палочки при поглощении фотона носит характер коллективного квантового явления, подобного усилению индуцированного излучения в активной среде лазера. Природа явлений может рассматриваться аналогичным образом: возникновение неравновесного, но стационарного состояния с накоплением значительной энергии, которая может высвобождаться при наличии фактора, выводящего систему из стационарности. Поскольку данное стационарное состояние не является равновесным и поддерживается только за счет значительных затрат энергии, высвобождаемая энергия на много порядков превышает энергию фактора, влияющего на выход из стационарности (в данном случае — поглощенного фотона). Запуск лавинообразного процесса высвобождения накопленной энергии возможен при условии соблюдения когерентности, т.е. сфазированности отдельных актов открытия ионных каналов. Поскольку этот процесс реально имеет место, следовательно, и условие когерентности соблюдается. Можно с определенностью утверждать, что поведение живой системы (в данном случае — органа зрения) управляется не классическими, а квантовыми законами, и изучение механизмов зрения с особой силой подчеркивает роль оптического диапазона электромагнитных волн как особого «мостика» между живой и неживой природой.
В настоящее время активно исследуются процессы регистрации света колбочками. Процессы в колбочках изучены пока недостаточно даже для того, чтобы хотя бы построить аналогичную палочкам самосогласованную модель. Установлено тем не менее с достаточной определенностью, что максимумы поглощения иодопсина и остальных опсинов распределены с достаточной степенью равномерности по всему видимому спектру (см. выше). Отсутствие хотя бы одного из опсинов вызывает искажение воспринимаемого спектра (дальтонизм). Зрение человека характеризуется наибольшим динамическим и наибольшим спектральным диапазоном из всех млекопитающих.
3.2.3. Фотосинтез как пример преобразования энергии биообъектами
В живой природе преобладают процессы энергообмена между электромагнитным излучением оптического диапазона и физико-химическими реакциями внутри биообъектов. Те организмы, которые синтезируют все необходимые им органические вещества за счёт поглощения энергии света, называются фототрофными ( или фототрофами), а те, которым для этого нужна химическая энергия — хемотрофными ( или хемотрофами).
Для фототрофов характерно наличие пигментов, которые поглощают энергию видимого излучения и превращают её в химическую энергию в ходе процесса фотосинтеза, на который затрачивается около 1% энергии излучения, достигнувшего поверхности Земли, что позволяет связывать около 75·1012 кг углерода в год. Фотосинтез в зеленых растениях определяет существование всех форм жизни, поскольку именно фотосинтез дает атмосферный кислород.
Уравнение фотосинтеза:
СО2 + Н2О  (СН2О) n + О2 + 470кДж/моль (3.5)
(СН2О) n + О2 + 470кДж/моль (3.5)
Кислород является отходом фотосинтеза. Полезный продукт — углеводы. Впрочем, в данном случае понятия «полезный продукт» и «отход» весьма относительны, поскольку кислород не менее необходим для существования биосферы, чем углеводы.
Спектром биологического действия[1] фотосинтеза называется зависимость скорости приращения количества кислорода N к от числа поглощённых квантов N и длины волны l, т.е. функцию  .
.
Если N к – число молекул кислорода, выделяемых в 1 с, N – число квантов, поглощаемых в 1 с, то  = hфс(l) – квантовый выход фотосинтеза. На рис. 3.9 и 3.10 показаны спектры действия фотосинтеза для зелёных растений и для хлореллы. Из рис. 3.9 видно, что спектр действия фотосинтеза практически совпадает со спектром поглощения хлорофилла. При выделении одной молекулы кислорода поглощается 8 квантов.
= hфс(l) – квантовый выход фотосинтеза. На рис. 3.9 и 3.10 показаны спектры действия фотосинтеза для зелёных растений и для хлореллы. Из рис. 3.9 видно, что спектр действия фотосинтеза практически совпадает со спектром поглощения хлорофилла. При выделении одной молекулы кислорода поглощается 8 квантов.
Скорость фотосинтеза зависит от интенсивности падающего света I=Nh ν. Для количественного определения скорости фотосинтеза можно воспользоваться эмпирической формулой:
 (3.6)
(3.6)
Здесь k ф– константа скорости ферментативной реакции (образования некоего субстрата из СО2 и Н2О при участии хлорофилла с последующим выделением О2). Субстрат образует слабо связанный комплекс, распадающийся на углеводные фрагменты и кислород. Для производства одной молекулы О2 требуется в среднем n = 8молекул субстрата, а на один ферментативный комплекс (неустойчивую молекулу субстрата) требуется от 50 до 300 молекул хлорофилла;  — концентрация фермента; К М — постоянная, имеющая смысл концентрации субстрата, при которой скорость реакции равна половине максимальной (константа Михаэлиса). При большой интенсивности падающего света I» K Мнаступает насыщение:
— концентрация фермента; К М — постоянная, имеющая смысл концентрации субстрата, при которой скорость реакции равна половине максимальной (константа Михаэлиса). При большой интенсивности падающего света I» K Мнаступает насыщение: 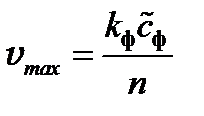 . Экспериментально k фможно определить, измеряя зависимость выхода О2 от продолжительности интервалов времени между короткими вспышками света. Эмпирически определено среднее время превращения одной молекулы субстрата: t d ≈ 0,02 с. Отсюда k ф ≈ td -1 ≈ 50 c-1 . Концентрация фермента для зеленых растений: = [Хл]/300 ([Хл] – концентрация хлорофилла). Отсюда
. Экспериментально k фможно определить, измеряя зависимость выхода О2 от продолжительности интервалов времени между короткими вспышками света. Эмпирически определено среднее время превращения одной молекулы субстрата: t d ≈ 0,02 с. Отсюда k ф ≈ td -1 ≈ 50 c-1 . Концентрация фермента для зеленых растений: = [Хл]/300 ([Хл] – концентрация хлорофилла). Отсюда
 . (3.7)
. (3.7)
Итак, максимальная скорость фотосинтеза при І = const равна одной молекуле кислорода на молекулу хлорофилла за 50 с.


У зелёных растений фотосинтез происходит в хлоропластах, которые рассеяны в цитоплазме. Они содержат хлорофилл и другие пигменты, локализованные в системе мембран, погруженных в основное вещество — строму. Световые реакции фотосинтеза происходят именно в системе мембран. В строме происходят темновые реакции.
Фотосинтетические пигменты высших растений — хлорофилл и каротиноиды. Их роль — поглощение света и превращение его энергии в химическую энергию. Различают формы хлорофилла — a, b, c, d; бактериохлорофилла — a, b, c, d, каротиноида — каротины, ксантофиллы (кислородсодержащие каротиноиды), b-каротин. Хлорофиллы поглощают главным образом красный и сине-фиолетовый свет (см. рис. 3.9). Зелёный свет они отражают, чем обусловлена зелёная окраска листьев большинства растений. Каждая форма отличается друг от друга по положению максимума поглощения в «красной» области оптического спектра: 670, 680, 690 и 700 нм.
 В темноте молекула хлорофилла находится на нижнем (основном) уровне S 0 (рис. 3.11). Полосы поглощения хлорофилла, соответствующие переходам S 0 →S* и S 0 →S 1 *, лежат в красной и синей областях спектра. Обратные переходы S * → S 0 и S 1 * → S 0 являются соответственно флуоресцентным и безызлучательным. Возможен конверсионный переход S *→ T. Уровень Т — метастабильный, поскольку излучательный переход Т→ S 0 запрещен по спину. Тем самым молекула хлорофилла долго пребывает в Т -состоянии, из которого возможно поглощение с переходом Т→Т*. Вследствие неспаренности электронов в Т -состояниях именно этот факт обуславливает возбуждение химических реакций.
В темноте молекула хлорофилла находится на нижнем (основном) уровне S 0 (рис. 3.11). Полосы поглощения хлорофилла, соответствующие переходам S 0 →S* и S 0 →S 1 *, лежат в красной и синей областях спектра. Обратные переходы S * → S 0 и S 1 * → S 0 являются соответственно флуоресцентным и безызлучательным. Возможен конверсионный переход S *→ T. Уровень Т — метастабильный, поскольку излучательный переход Т→ S 0 запрещен по спину. Тем самым молекула хлорофилла долго пребывает в Т -состоянии, из которого возможно поглощение с переходом Т→Т*. Вследствие неспаренности электронов в Т -состояниях именно этот факт обуславливает возбуждение химических реакций.
При поглощении света хлорофилл переводится в возбужденное состояние с потерей электрона (т.е. хлорофилл является донором электрона), который будет принят другой молекулой — акцептором электрона. Хлорофилл при этом окисляется, а акцептор восстанавливается.

 Световые реакции. Су-ществует два типа фотосин-тетических единиц (фотосис-темы I и II), которые состоят из набора молекул пигмен-тов, передающих энергию на одну молекулу главного пигмента – реакционный центр (РЦ) (рис. 3.12). В нём энергия используется для осуществления химических реакций. Здесь происходит преобразование световой энергии в химическую — главное событие фотосинтеза.
Световые реакции. Су-ществует два типа фотосин-тетических единиц (фотосис-темы I и II), которые состоят из набора молекул пигмен-тов, передающих энергию на одну молекулу главного пигмента – реакционный центр (РЦ) (рис. 3.12). В нём энергия используется для осуществления химических реакций. Здесь происходит преобразование световой энергии в химическую — главное событие фотосинтеза.
Оптимальная интенсивность фотосинтеза наблюдается при определенном соот-ношении между количеством возбуждённых фотосистем I и II (ФСI и ФСII), которое зависит от спектрального состава света. Свет поглощается обеими системами одновременно, но в разной степени. При смещении к крас-ной границе видимого спектра система ФСІ получает избыточную, а ФСІІ — недостаточную энергию. Баланс изменяется при смещении к синей границе. При изменении интенсивности падающего света энергия перераспределяется между ФСІ и ФСІІ.
Фотосистемы I и II взаимодействуют последовательно в так называемой Z–схеме фотосинтеза; восстановленные продукты ФСII служат донорами электронов для ФСI. Возбуждение светом, который в основном поглощается ФСII, приводит к восстановлению промежуточных переносчиков, а возбуждение ФСI — наоборот, к их окислению (рис. 3.13).
Поглощение света происходит пигментами светособирающего (СС) пигмент-белкового (ПБ) комплекса (ССПБК) фотосинтетических единиц, от которого, как из резервуара, энергия возбуждения передаётся на «антенный» ПБК ФСI и ФСII и далее непосредственно к реакционным центрам Р700 и Р680. Основной принцип работы реакционных центров состоит в том, что возбуждение фотоактивного пигмента приводит к отрыву от него электрона и восстановлению первичного акцептора, который потом передаёт электрон к следующему акцептору и далее – в электронно-транспортную цепь Z-схемы. При этом вероятность переноса электрона с возбуждённого уровня реакционного центра на первичный фотохимический акцептор намного выше, чем вероятность излучательной или безызлучательной диссипации энергии. Доставка энергии электрона возбуждения к реакционным центрам ФСI и ФСII высших растений и реакционному центру бактериального фотосинтеза осуществляется за счёт миграции энергии в светособирающей антенне путём безызлучательного переноса. Пигментные системы ФСI и ФСII пространственно разобщены, что затрудняет миграцию энергии электрона возбуждения между ними. Она осуществляется по механизмам индуктивного резонанса, а в пределах одного комплекса, где расстояния между молекулами пигментов составляют 0,9...1,4 нм, она носит экситонный характер. В таких системах делокализация экситона происходит за времена от 10-13 до 10-14 с.
Функциональное биологическое значение светособирающей «антенны» состоит в повышении эффективности использования поглощённых квантов. Среднее время, необходимое для утилизации энергии кванта света (выделение молекулы O2), составляет 0,01...0,02 с. Время регенерации фотоактивного пигмента реакционного центра намного меньше (£10-6 с). При ярком дневном свете каждая отдельная молекула Хл будет поглощать не более 10 квантов в секунду, а при обычной интенсивности света ещё меньше. В этом случае реакционный центр, практически лишённый собственной светособирающей антенны и включающий в свой состав фотоактивный пигмент, будет значительную часть времени «простаивать». Объединение многих десятков молекул пигментов в фотосинтетическую единицу, обслуживающую фотоактивный пигмент, предотвращает такого рода потери. Экспериментально показано, что время жизни (τ) синглетного возбуждённого состояния хлорофилла в растворе составляет 5 нс, а в хлоропластах оно сокращается в 20...30 раз (рис. 3.14). Очевидно, что при попадании возбуждения в реакционный центр происходит эффективный процесс использования электронной энергии состояния S 1 в первичном акте фотосинтеза. Тем самым время пребывания хлорофилла в возбуждённом состоянии S 1 в листе сокращается.
 Перенос электронов приводит к образованию АТФ[2] (фотофосфорилирование) и НАДФ·Н2:
Перенос электронов приводит к образованию АТФ[2] (фотофосфорилирование) и НАДФ·Н2:
 , (3.8)
, (3.8)

Таким образом, энергия света превращается в энергию химических связей АТФ и НАДФ·Н2. Вода расщепляется на Н2 и О2; Н2 переносится на НАДФ, а О2 — отход процесса фотосинтеза.
Темновые реакции происходят в строме и не требуют света. Их результатом является восстановление СО2 с образованием органических соединений – углеводов, при этом используется энергия химических связей АТФ и водород НАДФ·Н2.
3АТФ 3АДФ + Фн
СО2 + Н2О [СН2О] + Н2О (3.9)
2НАДФ · Н2 2НАДФ
Суммарное уравнение фотосинтеза:
2Н2О [CН2О]
Свет + Хлорофилл (3.10)
О2 2Н2 СО2 + Н2О
Таким образом, процесс фотосинтеза можно разбить на три этапа:
фотофизический – возбуждение электрона и миграция его энергии возбуждения (время от 10-16 до 10-12 с);
фотохимический – разделение зарядов, образование промежуточных продуктов (время от 10-12 до 10-2 с);
ферментативный – синтез биологических продуктов
(время от 10-2 до 104 с).
Первичные реакции фотосинтеза, начиная от поглощения кванта и кончая его захватом в реакционном центре (ферментативной реакции), протекают за время τ1 ≤ 100 нс. За это время не успевают полностью высветиться даже те переходы, которые не запрещены по спину. Поэтому квантовый выход первичного процесса велик (около 90 %). Вторичные процессы, приводящие к генерации О2, пока что не нашли однозначного и тем более количественного объяснения.
Другие фотосинтетические пигменты — каротиноиды (жёлтые, оранжевые, красные или коричневые) — сильно поглощают в сине-фиолетовой области. Они обычно замаскированы хлорофиллами, но выявляются перед листопадом, т.к. хлорофиллы разрушаются первыми. Каротиноиды содержаться также в цветах. Они функционируют как дополнительные пигменты. Каротиноид поглощает свет, переходя в метастабильное состояние и тем самым защищает хлорофилл от избытка света и окисления кислородом, выделяющимся при фотосинтезе. Защитная роль каротина определяется тем, что скорость его возбуждения и передачи энергии от возбужденной молекулы кислорода на 4 порядка выше, чем скорость окисления хлорофилла. Каротиноиды являются примером физического тушителя возбуждения (см. далее: роль β-каротина в преодолении побочных действий фотодинамического эффекта после сеанса фотодинамической терапии).
Молекулы каротиноидов, помимо светособирающей и защитной функции, выполняют роль стабилизаторов структуры пигмент-белковых комплексов фотосинтезирующих бактерий, высших растений и водорослей. Кроме того считается, что они повышают устойчивость микроорганизмов и водорослей к ионизирующему излучению, играют существенную роль в приспособлении к жизни в экстремальных условиях: сероводородной атмосфере, высокотемпературных источниках, концентрированных растворах солей и кислот.
3.2.4. Фотосенсибилизация. Принципы фотодинамической терапии (ФДТ).
Фотосенсибилизаторы. В медицине имеют большое значение фотопроцессы, возникающие в результате поглощения света экзогенными (внешними) веществами или эндогенными (внутренними) соединениями, концентрация которых резко возрастает при некоторых патологиях (протопорфирии, билирубинемии и др.). Вещества, повышающие чувствительность организма к УФ и видимому свету, называются фотосенсибилизаторами (Фс). По механизму действия они делятся на два типа. Фс первого типа (ФсI) под действием света сами химически изменяются, т.е. первичной стадией реакций первого типа служит образование свободных радикалов Фс. Эти реакции не зависят от присутствия кислорода О2. Например, более 4% населения болеет псориазом. Эта болезнь характеризуется гипертрофией эпидермиса, приводящей к покраснению и болезненному отшелушиванию кожи. Часть больных псориазом страдает также и псориастическим артритом. В терапии псориаза широко используют ФсI псорален. Фс второго типа (ФсII) — фотодинамические соединения. В реакциях второго типа первичным является взаимодействие возбуждённых молекул Фс с О2. Краткая схема реакции:
 (3.11)
(3.11)
ФсII, поглотив квант света, переходит в триплетное возбуждённое состояние (A T1), а затем взаимодействует с О2. При этом происходит передача энергии триплетного состояния ФсII молекулярному кислороду, благодаря чему он может переходить в возбуждённое синглетное состояние 1O2, в котором он примерно в 500 раз по сравнению с невозбуждённым O2 и в 100 раз по сравнению с кислородными и кислородсодержащими радикалами сильнее окисляет липиды, белки и другие биомолекулы.
В некоторых случаях ФсII образует супероксид–анионрадикал путём переноса на О2 электрона:
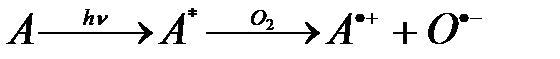 .
.
Реакции с ФсII, тем самым, существенно зависят от присутствия кислорода О2.
Типичный представитель ФсII — протопорфирин. Он накапливается в аномально высоких концентрациях в организме человека при некоторых нарушениях биосинтеза порфиринов. Если больные находятся под влиянием дневного (солнечного) света, то у них развивается эритема и эдема. При этом специальные тушители синглетного кислорода, т.е. вещества, принимающим на себя избыточную энергию возбуждения 1O2 и естественным образом присутствующие в клетке, не справляются со своей задачей, поэтому терапевтический метод лечения таких заболеваний — приём b-каротина (сильный тушитель-антиоксидант).
ФсI и ФсII широко используются в медицине, в том числе в фотодинамической терапии.
Физические основы фотодинамической терапии (ФДТ). ФДТ является относительно новым методом лечения злокачественных новообразований, получившим мощное развитие в течение последних 30 лет. Хотя такой срок является недостаточным с точки зрения всеобъемлющей оценки возможностей методики (в том числе и отдаленных результатов лечения), ФДТ уверенно заняла свое место рядом с традиционными методами, такими, как рентгено-, гамма- и химиотерапия. Обусловлено это тем, что традиционные методы лечения (в том числе и хирургические) во многом оказываются малоэффективными даже при сочетанном и комбинированном применении. Поэтому новый метод, отличающийся минимальной инвазивностью, практической безболезненностью и потенциально высокой эффективностью, связанной с селективностью воздействия на патологию, сразу же привлек пристальное внимание. В развитие метода вкладываются значительные силы и средства. Научно-лечебные центры, специализирующиеся на методе ФДТ, существуют более чем в 40 странах. Ведущими являются США, Германия, Япония. В последние годы значительные успехи достигнуты в Китае.
В России метод ФДТ известен давно, однако он долгое время не был разрешен к клиническому применению и вплоть до 1992 г. оставался на стадии исследований.

| Рис. 3.15. Структурная формула молекулы гематопорфирина IX. |
Основой химической структуры порфиринов является порфириновый цикл, состоящий из четырех пиррольных колец (рис. 3.15). Типы порфиринов различаются в зависимости от заместителей по 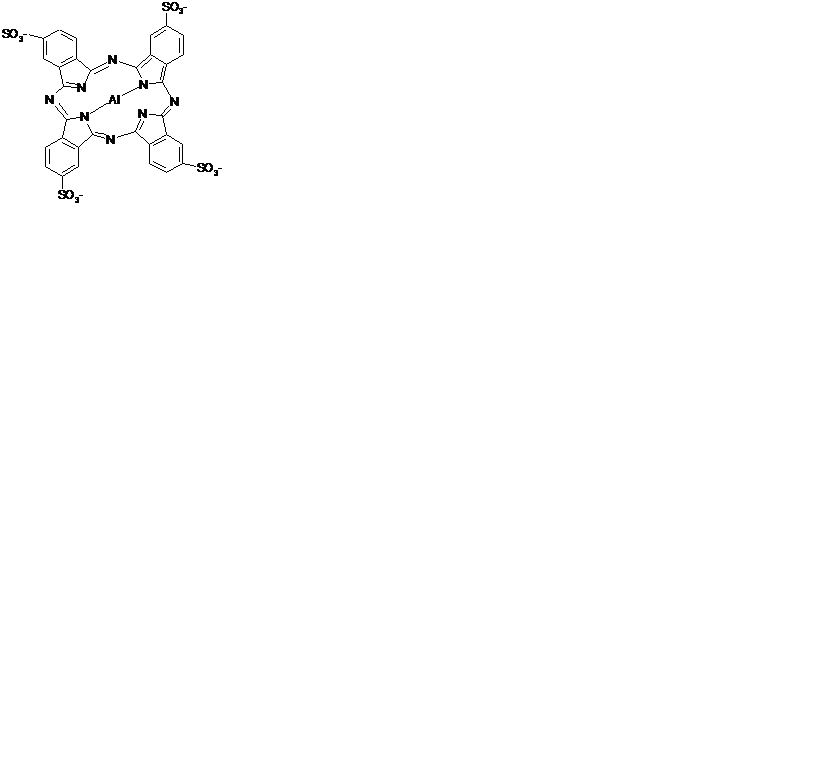
| Рис. 3.16. Структурная формула молекулы фталоцианина с атомом металла алюминия в центре. |
Эндогенные внутриклеточные порфирины существуют в виде металло-комплексов (с атомом металла в центре порфиринового кольца). Для ФДТ используют свободные порфирины (кроме фталоцианинов (рис. 3.16)), т.к. они обладают более низким коэффициентом тушения 1O2, чем металлокомплексы.
Все порфирины имеют сходные электронные спектры с четырьмя максимумами поглощения в видимой области и интенсивной полосой Сорэ с максимумом в районе 400 нм (рис. 3.17). Положение максимума полос зависит от растворителя.
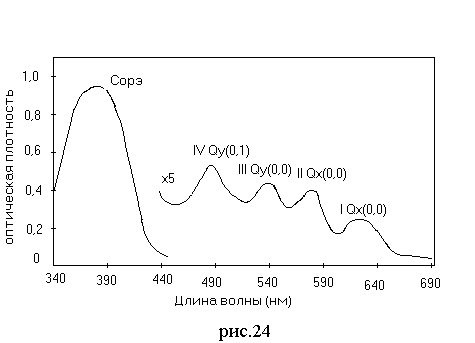
Рис. 3.17. Спектр поглощения серноуксускной ПГП.
При облучении тканей с накопленным Фс начинается фотохимическая реакция с синглетным кислородом 1O2, который вызывает повреждение клетки.
Квант света возбуждает первый синглетный уровень S 1 Фс, и далее происходят следующие реакции (рис. 3.18):
S 0 + h n  S 1 *; S 1 *
S 1 *; S 1 *  S 1 ; S 1
S 1 ; S 1  S 1 * + h n1 ; (3.12)
S 1 * + h n1 ; (3.12)

| Рис. 3.18. Схема передачи энергии возбуждения от фотосенсибилизатора к синглетному кислороду. |
S 1 *
 T 1 ; T 1
T 1 ; T 1  S 0 + h n2;
S 0 + h n2;
Здесь n1, n2 - частоты флуоресценции и фосфоресценции с возбуждённых уровней S 1 и Т 1 на основной уровень S 0; ki - константы скоростей перехода. Они обратно пропорциональны временам жизни на соответствующем уровне: k 0 ~ 1016 1/c; kk ~ 1012 1/c; kf ~ 108¸109 1/c; kq ~103¸104 1/c.
Для большинства порфиринов квантовый выход gик интерконверсии близок к единице:
 .
.
Следовательно, большая часть возбуждённых молекул с уровня S l переходит на уровень Т l, остальные дезактивируются в процессе флуоресценции. Излучательное время жизни уровня Т l (фосфоресценция) достаточно велико: t Т = 1/ kq = 10-3¸10-4 с, следовательно, возможен безызлучательный механизм передачи энергии другим молекулам, в частности кислороду О2. При этом О2 переходит на один из возбуждённых уровней, что подтверждается наличием характерных полос люминесценции кислорода.
Основное состояние молекулы кислорода 3О2 является триплетным, тогда как ближайшие возбужденные состояния синглетны (1О2*). Наиболее устойчи-вым из них является состояние 1D g, отстоящее от основного примерно на 0,976 эВ (l = 1,27 мкм). Ввиду различной мультиплетности состояний 1D g и основно-го 3О2 возможна аккумуляц

