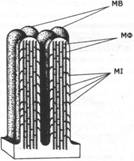
,.,.. - стаБМnИЭМРУlOщиА белОК ТРОПQМИО3ИН.
СБ - сшивающий белОК ПЛ - nлаэмалемма
РБ - режущиА бело.
ЯБ - якорн ••• А ёееск
КБ - кэпнрующмА белок

Поиск: Рекомендуем:  Почему я выбрал профессую экономиста
Почему я выбрал профессую экономиста
 Почему одни успешнее, чем другие
Почему одни успешнее, чем другие
 Периферийные устройства ЭВМ Периферийные устройства ЭВМ
 Нейроглия (или проще глия, глиальные клетки) Нейроглия (или проще глия, глиальные клетки)
Категории:
|
е) взаимодействие с маэмолеммоЯ *) РеryЛЯЦМJl полимеризации - деnonммермэацми.
,.,.. - стаБМnИЭМРУlOщиА белОК ТРОПQМИО3ИН. СБ - сшивающий белОК ПЛ - nлаэмалемма РБ - режущиА бело. ЯБ - якорн ••• А ёееск КБ - кэпнрующмА белок |
| Режущие белки, относящиеся к регуляторным АСБ, присоединяются к F-актину и вызывают его деполимеризацию или фрагментацию (разрезание длинной нити на несколько более коротких) (рис. 5д). Часть таких белков (гельзолин, виллин) не требуют для своей функции ионов Са2+, Т.е. являются Са1'-независuмыми. Они взаимодействуют с плюс-концом МФ, блокируя полимеризацию, и индуцируют фрагментацию МФ. Другая группа режущих белков (фрагмин, северин) представлена Са}' -зависимыми белками, активирующимися при определенной концентрации этого иона. Они также присоединяются к· плюс-концу и разрезают МФ на короткие фрагменты. Благодаря этому при значительных концентрациях Са2+ в периферической гиалоплазме возможна «взрывная» (очень быстрая) деполимеризация МФ. Большинство режуши.х белков - это мономеры с молекулярной массой 45-95 кДа). Однако обнаружены и низкомолекулярные представители этой группы, например, актиндеполимеризующий фактор (19 кДа). Этот белок связывает G-актин, после чего приобретает способность к фрагментации МФ. Актиновые МФ в аСС выполняют пространственноорганизующую функцию, являясь одним из элементов цитоскелета. В частности, они способны взаимодействовать с интегральными белками плазмалеммы и тем самым ограничивать их подвижность в БЛС. С помощью МФ, соединяющих мембранные белки, осуществляется определенная пространственная ориентация таких белков по отношению друг к другу в плазмалемме. Отдельные МФ в клетке могут взаимодействовать друг с другом и формировать сложные структуры. Такое взаимодействие осуществляется с помощью особой группы АСБ - сшивающих белков. К первой подгруппе сшивающих белков относят элонгирующие белки, структура и функции которых практически не изучены. Тем не менее, при определенных условиях они вызывают объединение коротких фрагментов F-актина в длинную МФ, Т.е. индуцируют быструю полимеризацию. Вторая подгруппа сшивающих белков (фасции, фимбрин, виллин и др.) обеспечивает образование пучков МФ. Они, как |
| правило, 'являются мономерными и соединяют МФ параллельно друг другу - формируют пучок МФ. В зависимости от строения белка, могут образовываться плотные пучки МФ (нити натяжения) (рис. 5б) или рыхлые пучки МФ (рис. 5в). Третья подгруппа сшивающих белков представлена гомодимерами (филамин; а-актинии, фодрин, актияогелин и др.), с помощью которых образуются сети МФ. эту подгруппу белков называют желактирующими белками, или желактинами. Данные белки каждым из своих протомеров взаимодействуют с одной МФ, объединяя их непараллельно, в результате чего формируются трехмерные сети МФ, расположенные в периферической гиалоплазме (рис. 5г). Еще одну группу АСБ, которую можно отнести к сшивающим белкам, представляют якорные белки. Функцией таких, в большинстве мономерных, белков (винкупин, талии, спектрин и др.) является прикрепление отдельных МФ, их пучков и сетей к белкам плазмалеммы (рис. 5е). Большинство якорных белков взаимодействует с внутренними участками МФ, однако, существуют и якорные белки, фиксирующие концы МФ или пучков МФ. Такие белки можно относить и к кэпирующим белкам. Действие сшивающих белков регулируется клеткой: они могут быть инактивированы либо путем расщепления с помощью ферментов протеаз, либо путем фосфорилирования с по- мощью протеинкиназ. Пучки И сети МФ, как и отдельные МФ, являются элементами цитоскелета в составе СОСА. Сети МФ могут выполнять и пространственно-организующую роль в отношении белков плазмалеммы, определяя их взаимное расположение в билипидном слое и ограничивая их подвижность. Пучки актиновых МФ, прикрепляясь концами к определенным белкам плазмалеммы, выполняют функции нитей натяжения, или стресс-фибрилл, обеспечивая противодействие осмотическому давлению, направленному на увеличение объема клетки. При разрезании нитей натяжения лазерным лучом происходит набухание (увеличение объема цитоплазмы) |
| 15 |
| клеток, которое может приводить к разрыву плазмалеммы и цитолuзу (разрушению клетки). Аналогичные последствия вызывает внеклеточная гипотоническая среда (раствор с низкой концентрацией солей). В этом случае в клетку через плазмалемму начинает поступать большое количество молекул воды, внутриклеточное давление на плазмалемму резко возрастает и сгресс-фибриллы не справляются со своей функцией. В клетках эпителия тонкой кишки пучки актиновых МФ являются скелетной основой микроворсинок, увеличивающих площадь поверхности этих клеток, которые осуществляют пристеночное пищеварение и всасывание его продуктов. МФ таких пучков соединены сши-вающими белками (виллин и фuмбрuн) И С помощью якорного белка соединены с мембранным белком участка плазмалеммы, входящего в состав микроворсинки (рис. 6). |
| Рисунок 6: |
|
|
| МI |
|
|
|
MI - миозин мв - никроворсинки МФ - ммкрофМбриллы Г - головка Ш - wарнирная часть Х - ХВОСТ Специализированные рецепторные клетки внутреннего уха, вОЛОСК06ые клетки, выполняющие функцию рецепции звуковых сигналов, имеют стереоцилии, содержащие пучок актиновых МФ, соединенный с участками плазмалеммы. С помощью стереоцилий осуществляется восприятие звуковых колебаний даже с очень низкой амплитудой (до 1 нм), Обнаружены химические вещества, влияющие на полимеризацию МФ. Например, метаболиты некоторых грибов цитохалазины (низкомолекулярные гетероциклические соединения), попав в клетку, взаимодействуют с молекулами свободного G-актина. Такой комплекс присоединяется к плюс-концу МФ, блокируя полимеризацию F-актина, что при водит к его деполимеризации (разрушению МФ). Циклический пептид фаллоидин (токсин гриба бледной поганки) взаимодействует сразу с F-актином, вызывая его суперстабилизацию. В результате МФ теряют способность к деполимеризации, т.е, к перестройке, необходимой в определенных ситуациях. Известны наследственные болезни, обусловленные дефектами АСБ. К ним относится один ИЗ видов миодистрофии (слабости скелетных мышц) - миодистро фия Дюшена. Причина этой болезни - дефицит или нарушение структуры якорного АСБ, названного дистрофином. К 8-13 годам больные данной миодистрофией теряют способность ходить, а в более позднем возрасте многие из них погибают от дыхательной или сердечной недостаточности на фоне респираторных инфекционных заболеваний. Дистрофин - полипептид, включающий 3685 аминокислотных остатков и формирующий 4 домена. Один из доменов имеет структуру, сходную с известным сшивающим АСБ а-актинином. Главной функцией дистрофина является прикрепление комплекса актиновых МФ к белкам плазмалеммы. В мышечных клетках этот белок необходим для фиксации миофибрилл (сократительных мышечных фибрилл) к плазмалемме мышечного волокна (сарколемме). Дефекты структуры дистрофина вызывают нарушение сократительной функции мышечных клеток, что и приводит К развитию симптомов миодистрофии. |
| Актиновые МФ в клетке являются не только элементаM~ цитоскелета, но и компонентами одной из универсальных двигательных внутриклеточных систем - актомиозиновой системы (АМС). В состав АМС входит также двигательный белок миозин. В настоящее время известны 3 варианта этого белка: миозин 1 (кодноголовый» миозин), миозин Il (кдвухголовый» миозин) и миозин V (сдвухголовый» миозин с V -образным стержнем). Миозин II (МlI) представляет собой гетерогексамер, молекулярная масса которого достигает 500 кДа. В молекуле имеется 3 разных типа цепей (полипептидов): тяжелая цепь (ТЦ), легкая структурную цепь (ЛСЦ) и легкая регуляторная цепь (ЛРЦ). Таким образом, в молекуле MII присутствуют по две одинаковых ТЦ, ЛСЦ и лрц. ТЦ (1200 аминокислотных остатков) имеет 2 домена: глобулярный (головку), включающий чуть больше 800 аминокислотных остатков, и фибриллярный (стержень), длина которого составляет 150 нм, а толщина - 2 нм. Глобулярная ЛСЦ взаимодействует с ТЦ в проксимальной области головки, а ЛРЦ - на границе головки и стержня. Молекулярная масса каждой из ЛЦ составляет около 20 кДа. При образовании МJI 3 разных полипептида формируют гетеротример с крупной глобулярной головкой и длинным фибриллярным стержнем (рис. 7). Стержень такого тримера является п-спирапьным, благодаря чему стержневые участки двух одинаковых тримеров взаимодействуют друг с другом. В результате этого формируется молекула MII, состоящая из двух головок и общего двухспирального стержня (рис.7а). в жестком стержне МII имеются 2 гибких (шарнирных) участка - в середине стержня и на его границе с головкой. Они обеспечивают изменение положения головки по отношению к стержню, а также сгибание стержня в его центральном участке. Главная функция стержня - формирование миозиновых филаментов. В немышечных клетках и клетках гладких мышц образуются тонкие миозиновые фила менты путем взаимодействия двух молекул MII своими дистальными участками стержня по принципу «хвост к хвосту». В результате этого формируется тонкий миозиновый филамент, на обоих концах которого находятся двойные головки (рис. 7б). В клетках исчерченных мышц (скелетных и сердечной) обнаруживаются более длинные толстые миозиновые филаменты. Они состоят из большого количества молекул MII, взаимодействующих друг с другом двумя способами (рис. 7в). Первый из них - антипараллельный, характерный для образования и тонких миозиновых филаментовЛри втором способе молекулы МII объединяются параллельно друг другу различными районами дистальной части стержня. В результате таких взаимодействий формируется биполярный толстый миозиновый филамент. В центре филамента расположена ЗОI1а, не имеющая головок, а на концах - две зоны, на поверхности которых спирально расгюложены многочисленные (около 500) двойные миозиновые головки. Функции миозиновых головок обусловлены наличием в каждой из них А Тфазного центра и нескольких актинсеязывающих центров. В АТФазном центре осуществляется присоединение и гидролиз А ТФ (А ТФазная реакция, которая является Mg2+-зависимоЙ). В ходе АТФазной реакции происходит изменение конформации головки, в результате чего изменяется ее сродство к F-актину и положение по отношению к стержню, что позволяет головке «шагать» по МФ (рис. 8). Находясь в комплексе с А ТФ, головка миозина не обладает сродством к F-актину. Гидролиз АТФ приводит к тому, что вАТФазном центре оказывается комплекс АДФ·Ф, (неорганический фосфат), вызывающий изменение конформации головки. Новое конформационное состояние головки характеризуется активацией ее актинсвязывающих центров, в результате чего головка взаимодействует с F-актином. Связывание головки с F-актином при водит к дальнейшему изменению ее конформации и выводу Ф, из А ТФазного центра. Следствием этого является формирование сильных связей между головкой миозина и r-актином, изменяющее положение головки по отношению к стержню миозина (осуществление рабочего хода головки). Таким образом, рабочий ход головки обеспечивает движение молекулы миозина по F-актину. Новый цикл работы АМС начинается с замены АДФ в головке миозина на молекулу АТФ. это событие приводит к изменению конформации головки, потере ее связи с F-актином и возвращению в исходное положение по отношению к стержню. В результате головка миозина после гидролиза А ТФ получает возможность взаимодействовать с другим участком F-актина и делать очередной шаг по МФ в направлении от ее минус-конца к плюс-концу. |
| 16 |
| PUCVIlOK 7: |
| В) миозин n (МII) и его про_с. а) Crpoeииc IЮDel<)'JIJoI МII б) ТoвпII МW(I1IПIO •••• фllJ8МeШ В) ТoлcndIlOЮЗIIВOIIwJI фlDlUlсиr С - crep:aсиь Ш - шapивpвu оБJIIICIЬ Г - I'0JJ08J<В |
| PUCVIlOK 8: |
| МФ |
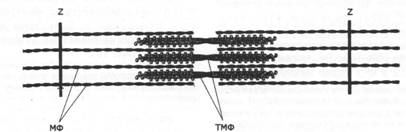
| PUCVIIOK 9: |
| Строение саркомера МФ - МИКРОфибриллы ТМФ - толстые миозиновые филаменты. Z - Z диск Реально в состав АМС входят не отдельные молекулы MII, э миозиновые филаменты (тонкие или толстые) с головками на обоих концах - биполярные миозиновые филаменты. С одЧОЙ стороны, это дает возможность взаимодействия с двумя или более) параллельно расположенными МФ. Однако, с дру-ой стороны, головки таких миозиновых филаментов должны шагать» по МФ в разных направлениях. В результате этого происходит движение не миозиновых C'Lla.\1eHTOB по МФ, а МФ по отношению к миозиновому ~:. "-lа~tенту, причем разные МФ перемещаются в противопо_-~жных направлениях. Таким образом, миозиновые фила~ енгы в АМС обеспечивает взаимное скольжение МФ, СВЯt::..-"'!bl.\ с головками разных концов миозинового филамента. Ес..1И концы МФ прикреплены к белкам плазмалеммы, .. ~ -\.\1С приводит к их сближению в БЛС слое, что .... _:;~ нкциональное значение для определенных мем -: - - -;х белов. Когда мембранные белки или белковые.:.....~~cы зафиксированы в плазмалемме достаточно же- |
|
|
| Схема работы актомиозиновоА системы М - миозин МФ - микрофибрилла Фн - неорганический фосфат |
|
|
|
|
| стко, действие АМС приводит к сокращению клетки, что является важнейшей функцией мышечных клеток. В клетках исчерченной мускулатуры функциональной единицей АМС, обеспечивающей процесс сокращения, является саркомер (рис. 9). Его границы формируются поперечно расположенными белковыми Z-дuскамu. От каждого из этих дисков навстречу друг другу отходят многочисленные МФ, которые в скелетной мускулатуре прикрепляются к дискам молекулами фибриллярного белка небулина, расположенными вдоль МФ. В клетках сердечной мышцы небулин отсутствует. МФ прикреплены и к сложному гликопротеину плазмалеммы с помощью белка дистрофина. Толстые миозиновые филаменты расположены между МФ и прикрепляются одновременно к обоим дискам специальным белком коннексином (тайтином). Наследственные дефекты дистрофина при водят к развитию миодистрофий - болезней, характеризую- |
| 17 |
|
|
|
|
Дата добавления: 2018-11-11; Мы поможем в написании ваших работ!; просмотров: 166 | Нарушение авторских прав
Лучшие изречения:
 | 3927 -
| 3927 -