




Дисоціація молекули.
Реакції приєднання.
5. Реакції димеризації й приєднання (приводять до утворення кінцевих стабільних продуктів).
6. Реакції диспропорціонування (приводять до утворення кінцевих стабільних продуктів).
Реакції приєднання кисню.
Реакції перенесення водню.
Основною кількісною характеристикою радіаційно-хімічних реакцій є вихід продуктів реакцій G на одиницю поглинутої дози, тобто число молекул, що виникли внаслідок перетворень речовин у ході радіаційно-хімічних реакцій, на 100 еВ поглинутої енергії випромінювань.
Радіаційно-хімічний вихід G обчислюють так:
G = M / E,
де М – кількість змінених молекул; Е – поглинута енергія випромінювання, Е=100 еВ.
Іонний вихід:
І = М / N,
де N – число іонізацій.
Оскільки на акт іонізації витрачається енергія 32,5 еВ, то Gmax не може перевищувати значення 3, бо 100 еВ / 32,5 еВ=3. В СІ ця величина визначається як число молекул реакції на дозу 1 Гр.
Зазвичай при символі G у вигляді індексу вказують продукти відповідної радіаційно-хімічної реакції. Наприклад, для продуктів радіолізу води обчислюють виходи реакцій GH, GOH, GOH2, GO2. Сумарний вихід радіолізу води становить близько 4.
Величина Gx не є сталою, бо її значення залежить від дози опромінення й концентрації речовини, перетворення якої в радіаційно-хімічній реакції дає відповідний продукт х. Зі збільшенням дози опромінення, як правило, зростає й кількість перетворюваних молекул. Залежність виходу реакції від дози описується експоненціальними функціями. Характеристики цих експонент дають змогу кількісно описувати ефективність радіаційно-хімічних реакцій і визначати поперечний переріз реакції  – коефіцієнт при показнику експоненціальної функції.
– коефіцієнт при показнику експоненціальної функції.
Якщо кількість перетворюваних у радіаційно-хімічній реакції молекул позначити N, то
N = N0 * e- 
де N0 – початкова кількість молекул; D – доза опромінення; - поперечнийпереріз реакції.
Основну масу будь-якої живої клітини становить вода, тому і ймовірність влучання іонізуючого випромінювання в молекули води є високою. Відповідно значну роль у радіаційно-хімічних перетвореннях речовин у клітинах можуть відігравати продукти радіолізу води.
Унаслідок дії іонізуючого випромінювання на чисту нейтральну воду утворюються нестабільні продукти радіолізу, які зазнають подальшого перетворення, формуючи різні активні хімічні форми.
Первинними продуктами взаємодії випромінювання й води є іонізовані або збуджені молекули:
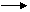 Н2О Н2О++е-,
Н2О Н2О++е-,
Н2О+Е Н2О*.
Радіаційно-хімічний вихід реакцій для розчинених речовин залежить від їхньої концентрації. Як правило, зі збільшенням концентрації речовини G зростає й досягає плато, що відповідає широкому діапазонові високих концентрацій. Цю залежність виходу радіаційно-хімічних реакцій від концентрації речовини було вперше описано В.Дейлом, який назвав її ефектом розведення.
Ушкодження ДНК, з одного боку, можуть бути пов’язані з розривом у каркасі молекул. Це може спричинятися розривом глікозильного зв’язку – між першим вуглецевим атомом пентоди та першим атомом піримідину або дев’ятим атомом азоту пурину. З іншого боку, можуть зазнавати хімічних змін азотисті основи, що супроводжується локальними порушеннями структури подвійної спіралі.
Унаслідок опромінення нуклеїнових кислот у водних розчинах або в сухому стані відбуваються радіаційно-хімічні перетворення як азотистих основ, так і вуглеводної частини молекули.
Унаслідок опромінення ДНК вільні основи й продукти їх радіаційно-хімічних перетворень частково відокремлюються від полінуклеотидного ланцюга у вигляді дрібних фрагментів, частково ж модифіковані основи залишаються в макромолекулі. З цього стану їх можна вивільнити відповідним гідролізом.
Процеси, які відбуваються внаслідок опромінення ДНК безпосередньо в живій клітині, ускладнюються тим, що до власне радіаційно-хімічних перетворень азотистих основ і розривів фосфоефірних зв’язків приєднуються вторинні реакції, в котрих беруть участь ферменти.
Зміни концентрації тих чи інших метаболітів або будь-яких інших речовин в опромінених клітинах мають різну природу. Такі зміни віддзеркалюють як прямі наслідки опромінення, так і далекі опосередковані ефекти, пов’язані з оз регулюванням гомеостатичних станів організму, ушкодженням мембран, імонологічними процесами тощо (рис. 1).
Біохімічні зміни, які відбуваються в організмах під впливом опромінення, є наслідком не стільки безпосередніх радіаційно-хімічних реакцій тих або інших речовин, скільки метаболічних змін в опромінених клітинах і тканинах. Такі зміни зумовлені тими функціональними порушеннями, які властиві опроміненим клітинам і організму в цілому.
Аналіз індукованих опроміненням процесів свідчить: поштовхом до їх початку є радіаційно-хімічні зміни певних речовин і зумовлені цими змінами функціональні порушення клітинних процесів – зміни швидкостей окремих метаболічних реакцій, зміни мембранної проникності, ушкодження окремих клітин, із чим може бути пов’язана поява токсичних речовин, які виникають унаслідок цитолітичних процесів, і т.ін. Унаслідок цих процесів, які іноді супроводжуються сильними зсувами в стані метаболічних систем клітин і швидкостей біогенезу різних речовин, можуть спостерігатися сильні зміни речовинного складу тканин, активностей окремих ферментних систем.
Характерним для опромінених тканин є більш або менш різкі зміни темпів біосинтезу окремих ферментів, зокрема, тих, що контролюють синтез ДНК. При цьому під впливом іонізуючого випромінювання синтез деяких ферментів прискорюється, тоді, як синтез ферментів, що контролюють синтез гормонів, гальмується.
Розкриття закономірностей біохімічних змін в опромінених організмів відображує динамічну інтегральну картину формування радіаційного ураження на рівні цілісного організму.
 НАСДІКИ ОПРОМІНЕННЯ
НАСДІКИ ОПРОМІНЕННЯ
Рис. 1
Регенерація органів, які перебувають в стані спокою. Верхівкова меристема пагона вищих рослин, яка активно росте, в значній мірі, а іноді і повністю, придушує поділ клітин в меристемах бокових бруньок. Це явище, що отримало назву апікального домінування, являє собою сильну міжклітинну взаємодію, яка відбувається на рівні цілісного організму. Видалення верхівки приводить до зняття апікального домінування і посилення проліферативної активності тканин, які перебувають в стані спокою і пробудженню бокових бруньок. Всі ці явища належать до регенераційних.
Апікальне домінування визначається гормональним типом впливу на точки росту. Це підтверджується чисельними дослідженнями, в яких за допомогою таких рістактивуючих речовин, як кінетин і гетероауксин, вдавалося знімати апікальне домінування або, навпаки, відновлювати його при видаленій верхівковій меристемі.
Опромінення рослин іонізуючою радіацією, природно, веде до більшого пошкодження верхівкових меристем, клітини яких активно діляться і ростуть. І якщо у рослини існують органи, які перебувають в стані спокою і мають підвищену радіостійкість, то її загибель повинна викликати їх регенерацію. І, безумовно, описані в радіобіологічній літературі факти, які свідчать про сильне гілкування опромінених рослин (дихотомія, фасціації), активацію утворення бокових коренів, про підвищення кущування та інше, пояснюється саме зняттям апікального домінування і посиленням регенераційних процесів.
Найкраще апікальне домінування вивчене у бобових рослин, які здавна вважаються класичними об'єктами для його демонстрації. Бокові бруньки у бобових розміщені в пазухах листків і тому називаються пазушними. За нормальних умов росту і розвитку вони знаходяться в стані спокою протягом всього онтогенезу. Але при знятті апікального домінування, наприклад, шляхом механічного видалення верхівкової бруньки, одна (іноді дві) бокова брунька вступає в ріст і розвивається в пагін, який повністю повторює організацію головного пагона і формує в подальшому всю рослину.
Аналогічна ситуація виникає при опроміненні рослин, як, до речі, і при пошкодженні апікальної бруньки хімічними агентами, її усиханні, відмерзанні і дії інших чинників. Існує мінімальне порогове значення дози опромінення, при якій апікальне домінування повністю знімається. Безумовно, воно залежить від видової радіочутливості рослини і визначається тим рівнем дози. який веде до припинення поділу у певної критичної кількості клітин і інгібування синтезу фітогормонів, що пригнічують ріст пазушних бруньок (рис. 7.14). Саме тоді клітини меристем пазушних бруньок приступають до проліферації.

Рис. 7.14. Взаємодія фітогормонів в рослині в нормі і при знятті апікального домінування в результаті опромінення іонізуючою радіацією:
1 - рослина в нормі: цитокініни (ц), що синтезуються в корені, транспортуються в апікальну бруньку пагона, викликаючи поділ її клітин, а синтезовані в ній ауксини (а), рухаючись вниз по стеблу, за допомогою індукції утворення етилену та абсцизової кислоти пригнічують поділ клітин бокових бруньок і центру спокою кореня; 2 - при опроміненні і знятті апікального домінування пригнічується синтез ауксинів, а цитокініни спрямовуються до бокових бруньок і центру спокою, клітини яких починають поділ; 3 - в пагоні відновлюється домінування нового апікального органу, а центр спокою відновлює меристему кореня.
Регенераційне відновлення дійсно слід віднести до найпотужнішої системи відновлення, якою еволюція нагородила вищі рослини, котрі не мають можливості захищатися від пошкоджень за допомогою інших засобів. З його основі лежать неспецифічні пристосувальні реакції, які розширюють межі зміни умов життя рослин, і механізм регенераційного відновлення ефективний при дії будь-якого пошкоджуючого агента.
Компенсаторне відновлення
Цей тип післярадіаційного відновлення є найменш вивченим в радіобіології, хоча компенсаторне посилення окремих функції у опромінених іонізуючими випромінюваннями організмів доводиться спостерігати нерідко. Серед них можна виділити два основних типи компенсаторних реакцій. Перший з них зв'язаний з виконанням непошкодженими внаслідок опромінення клітинами, тканинами, органами не властивих їм функцій. Другий - з посиленням функцій, коли непошкоджені клітини, тканини, органи посилюють власне навантаження з метою надолуження функції інших, які їх втратили.
Дедиференціація спеціалізованих клітин і тканин. У перебігу розвитку організму в ньому відбувається формування спеціалізованих клітин, тканин і органів за рахунок виникнення протягом морфогенезу відмінностей між однорідними клітинами і тканинами - процес диференціації. Так, меристематична клітина після здійснення декількох (п'яти-семи) поділів переходить до розтягнення, в процесі якого змінюється її форма і збільшуються розміри, потім до диференціації, при якій зовнішньо однакові клітини набувають різних властивостей та формують різні типи тканин, і далі до спеціалізації, яка характеризується різними рівнями метаболічної активності і ступенем структурної організації тканин і органів.
Звичайно, диференціація необоротна. Але в умовах пошкодження тканин, при злоякісному рості і деяких інших ситуаціях, що ведуть до зміни балансу фізіолого-активних речовин, природний хід руху клітин по життєвому циклу порушується, і вони можуть набути здатності до диференціації в зворотному напрямі, тобто дедиференціації. Саме такі умови можуть скластися при опроміненні рослин, коли, як згадувалося, змінюється баланс фітогормонів, і клітини меристеми втрачають здатність до поділу. При цьому диференційовані клітини починають ділитися і стають джерелом відновлення меристеми або виникнення нових твірних тканин.
Дедиференціація – один з найяскравіших прикладів компенсаторного відновлення, коли опромінені клітини набувають не властивих їм функцій.
До компенсаторного шляху післярадіаційного відновлення слід віднести нерідко констатоване після опроміненні посилення функцій клітин, тканин і органів, які зберегли свої функції. Так, при опроміненні кущових форм злаків при зменшенні кількості стебел ті, що вижили, як правило, бувають більш рослими і міцнішими, ніж окремі стебла в неопроміненому контролі. При пошкодженні генеративних органів і зменшенні кількості зав'язі на рослині сформовані колоски, стручки, плоди виявляються більш продуктивними, ніж в контролі. При зменшенні кількості зерен в суцвітті ті, що залишились, за абсолютною масою перевищують контрольні.
Ці факти можна пояснити з позицій зміни характеру перерозподілу поживних речовин, що надходять з кореневої системи - те, що призначалося для семи, дістається одному. В цьому відношенні не менш ілюстративними, але однозначно трактованими з позицій компенсаторного посилення функцій, є дані, одержані на рівні клітин, тканин. Зокрема, вище згадувалось, що при γ-опроміненні рослин в дозах, при яких кількість клітин в меристемах зменшується в 2-3 рази, різко зростає швидкість їх поділу. Виявляється, що клітини, які залишаються непошкодженими або відновили здатність до поділу, намагаючись поновити початковий клітинний об'єм меристеми, починають активно розмножуватися. В цей період після короткочасного гальмування швидкість поділу клітин суттєво зростала - майже на чверть скорочувалась тривалість їх клітинного циклу. Потім, по мірі досягнення меристемою норми, вона поступово збільшувалась і досягала початкового рівня (рис. 7.11).
Таке, навіть короткочасне, але досить суттєве посилення швидкості поділу клітин є характерним прикладом прояву компенсаторних явищ на рівні клітинної популяції.
Свідченням компенсаторного типу відновлення може бути нерідко спостережуване після опромінення у пошкоджуючих дозах тимчасове посилення синтезу нуклеїнових кислот, білків, фітогормонів, ферментів, макроергічних та інших сполук. Саме наслідком всіх цих процесів і є посилення поділу клітин, яке можна вважати їх інтегральним показником.
В цілому, розуміючі під терміном "відновлення” сукупність процесів, що забезпечують збереження організму від пошкодження або загибелі при дії іонізуючої радіації як системи, якій притаманне виконання певних функцій, варто уважно вивчати особливості прояву певних його реакцій, спрямованих на відновлення саме втрачених функцій. Управляючи цими процесами, можна сприяти більш або менш значному відновленню опромінених організмів у післярадіаційний період.

