Поиск:
Рекомендуем:
 Почему я выбрал профессую экономиста
Почему я выбрал профессую экономиста
 Почему одни успешнее, чем другие
Почему одни успешнее, чем другие
 Периферийные устройства ЭВМ
Периферийные устройства ЭВМ
 Нейроглия (или проще глия, глиальные клетки)
Нейроглия (или проще глия, глиальные клетки)
Категории:
Микротрубочки, реснички и центриоли. 2 страница
|
|
|
|
|
Поиск: Рекомендуем:
Почему я выбрал профессую экономиста
Почему одни успешнее, чем другие
Периферийные устройства ЭВМ
Нейроглия (или проще глия, глиальные клетки)
Категории:
|
Микротрубочки, реснички и центриоли. 2 страница
|
|
|
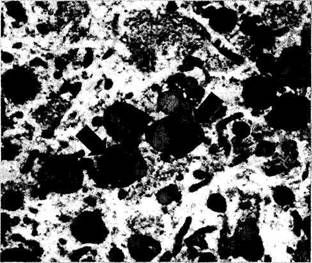
меланоцитах увеального тракта (рис. 1.1.23). Пигментные гранулы перечисленных двух оболочек довольно трудно различать при световой микроскопии. Гранулы, выявляемые в нейро-эпителии, несколько больше в размерах и темнее, чем обнаруживаемые в увеальном тракте. Второй тип гранул — липофусцин — может быть спутан с меланином, поскольку он имеет светло-коричневый цвет и по размерам приближается к зернам меланина (рис. 1.1.26). Гранулы липофусцина имеют более светлый цвет. Липофусцин чаще обнаруживается в пожилом и старческом возрасте, особенно в клетках миокарда и нейронах. Довольно много появляется его с возрастом в клетках пигментного эпителия сетчатки.
Рис. 1.1.26. Зерна липофусцина (стрелки) в цитоплазме нейрона (трансмиссионная электронная микро скопия)
В клетках нередко обнаруживаются и другие пигменты, такие как гемоглобин, билирубин. Продукты деградации гемоглобина, депозиты железа чаще обнаруживаются в патологически измененных тканях глаза (стекловидное тело, увеальный тракт).
Интерфазное ядро
Все клетки содержат ядра, форма и размеры которых могут быть самыми разнообразными. В настоящем разделе мы остановимся на морфологии интерфазного ядра, т. е. ядра, находящегося вне митотического цикла. В тканях подавляющее большинство ядер находится в интерфазе.
В ядре четко выявляются следующие структуры (рис. 1.1.27):
1. Ядерная оболочка.
2. Хроматин.
3. Ядрышко.
4. Ядерный сок.
Рис. 1.1.27. Ультраструктурные особенности лимфоцита периферической крови. Четко определяется структура ядра:
/ — эухроматин; 2 — гетерохроматин; 3 — митохондрии; 4 — ядро; 5 — ядрышко; 6 — ядерная оболочка; 7 — ядерная пора; 8 — плазмолемма; 9 — перинуклеарное пространство; 10 — псевдоподии; // — полирибосомы; 12 — шероховатый эндоплазмати-ческий ретикулум
Ядерная оболочка (кариолемма). Ядерная оболочка окружает ядро и хорошо видна в препаратах. Это связано с тем, что с внутренней стороны к ней прилежит хроматин, интенсивно окрашивающийся гематоксилином. Окрашенный хроматин и контурирует оболочку.
При исследовании в электронном микроскопе оболочка выглядит в виде двух темных мембран, между которыми определяется светлое пространство толщиной 25 нм. Толщина каждой электронноплотной мембраны 8 нм.
В ядерной мембране определяются многочисленные «поры» (рис. 1.1.28). Ядерные поры занимают от 3 до 35% всей поверхности ядра. Именно в этих местах два электронноплотных слоя оболочки как бы сливаются. В области пор обнаруживается скопление хроматина. Ультраструктурные исследования выявили, что в области пор располагаются и довольно сложные структуры, состоящие из канальцев, обращенных как в сторону цитоплазмы, так и внутрь ядра. Этот комплекс структур называют паровым комплексом (рис. 1.1.29). Поровый комплекс содержит два параллельных кольца (по одному с каждой поверхности кариолеммы) диаметром 80 нм, которые образованы 8 белковыми гранулами. От этих гранул к центру сходятся фибриллы, формирующие перегородку (диафрагму) толщиной 5 нм. В середине этой мембраны лежит центральная гранула, которая представляет собой субъединицу рибосомы. Поры способствуют обмену метаболитов между ядром и цитоплазмой. Основной функцией ядерных пор является обеспечение регуляции
Клетка
19

|
|
|
Рис. 1.1.28. Ультраструктурные особенности (а) и схематическое изображение (б) строения поры ядерной оболочки:
/ — пора; 2 — ядро; 3 — цитоплазма клетки; 4 — внутренняя мембрана ядерной оболочки; 5 — наружная мембрана ядерной
оболочки; 6 — гранулярный эндоплазматический ретикулум
|
|
Рис. 1.1.29. Поры ядерной оболочки:
а — сканирующая электронная микроскопия (стрелкой указан комплекс ядерной поры); 6 —ядерные поры при применении метода
замораживания-скалывания; в — схема организации комплекса ядерной поры (/ — наружная мембрана кариолеммы; 2 — внутренняя
мембрана кариолеммы; 3 — белковые гранулы; 4 — белковые фибриллы; 5 — центральная гранула)
избирательного транспорта веществ между цитоплазмой и ядром, активный перенос в ядро белков, перенос в цитоплазму субъединиц рибосом.
Хроматин при световой микроскопии выглядит в виде глыбок различной степени дисперсности, равномерно или неравномерно распределенных в кариоплазме (рис. 1.1.30). Отмечается его большая конденсация вблизи ядерной мембраны (периферический хроматин) и вблизи ядрышка (околоядрышковыи хроматин). Описанный тип хроматина называется конден сированным, поскольку он виден при световой микроскопии. Но кроме конденсированно-
го гетерохроматина в кариоплазме существует и неконденсированный хроматин, или эухро- матин.
В химическом отношении хроматин представляет собой комплекс ДНК и белка. Этот комплекс соответствует хромосомам, которые в интерфазном ядре представлены длинными, тонкими перекрученными нитями. Они неразличимы как индивидуальные структуры.
Транскрипция информации с молекул ДНК осуществляется только с молекул эухроматина.
Морфологической разновидностью гетерохроматина является тельце Барра, обычно расположенное вблизи ядерной оболочки. Обнару-
20
Глава 1. КЛЕТКА И ТКАНИ

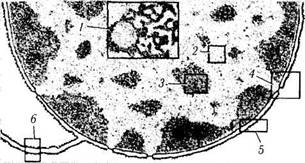
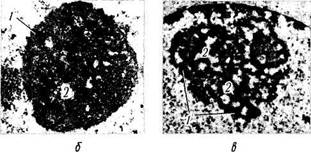
Рис. 1.1.30. Ультраструктурная организация ядра:
а — схема структурных компонентов ядра (/ — ядрышко; 2 — эухроматин; 3 — гетерохроматин; 4 — ядерная пора; 5 — карио-лемма; 6 — цистерна шероховатой эндоплазматической сети); б, в — особенности строения ядрышка (/—темный компонент; 2 — светлый компонент)
живаются тельца только у женщин, поскольку представляют собой одну из конденсированных Х-хромосом.
Функция реализации генетической информации в интерфазном ядре осуществляется непрерывно благодаря процессам транскрипции. При транскрипции ДНК образуется одна очень крупная молекула РНК (первичный транскрипт), которая связывается с ядерными белками с образованием рибонуклеопротеидов. В первичном РНК-транскрипте (как и в матричной ДНК) имеются дискретные значащие последовательности нуклеотидов (экзоны), разделенные длинными вставками (нитронами). Про-цессинг РНК-транскрипта включает отщепление интронов и стыковку экзонов — сплайсинг. При этом очень крупная молекула РНК превращается в достаточно мелкие молекулы информационной РНК, отделяющиеся от связанных с ними белков при переносе в цитоплазму.
Ядрышко (рис. 1.1.30). Ядрышко представляет собой расположенное в ядре плотное образование. Размеры, плотность, форма и локализация ядрышек могут быть самыми разнообразными. Отмечено, что более интенсивна синтетическая деятельность клетки при больших размерах ядрышка. Да это и понятно, поскольку ядрышко обеспечивает синтез РНК-
Ультраструктурные исследования позволили выявить довольно сложную структуру ядрышка. В нем различают гранулярный, фибриллярный и аморфный компоненты.
Гранулярный компонент представлен зернами (диаметр 10—20 нм), состоящими из ри-бонуклеопротеидных частиц (субъединицы рибосом). Фибриллярная часть состоит из плотных тонких электронноплотных нитей (диаметр 5—8 нм), образующих компактную массу. Эти волокна концентрируются вокруг более светлых сердцевин из менее плотного материала (фибриллярные центры). Считается, что фибриллярный материал представляет собой РНК (рибосомальная РНК), а фибриллярные центры состоят из ДНК и по строению соответствуют зернам хроматина.
Аморфный компонент окрашивается бледно и содержит участки расположения ядрышковых организаторов со специфическими РНК-связы-вающими белками и крупными петлями ДНК, активно участвующими в транскрипции рибосо-мальной РНК-Фибриллярный и гранулярный компоненты образуют ядрышковую нить (нуклеонему), толщина которой 60—80 нм.
Ядерный сок (кариоплазма). Ядерный сок представляет коллоидный раствор белка, в котором и располагаются перечисленные структуры. Ядерный сок не окрашивается ядерными красителями.
Основными функциями ядра является хранение генетической информации (в молекулах ДНК, находящихся в хромосомах), реализации генетической информации, контролирующей осуществление различных жизненных функций клетки, воспроизведение и передачу генетической информации. Последняя функция осуществляется благодаря клеточному делению.
Клеточное деление
Задачей этого раздела не является изложение основ генетики. Мы опишем лишь морфологические проявления деления клеток.
Необходимо отметить, что биологический смысл клеточного деления сводится к воспроизведению дочерней клетки, идентичной материнской. В генетическом плане суть деления сводится к тому, что материнская клетка, обладающая определенным набором хромосом (23 пары хромосом у человека), первоначально удваивает генетический материал, и только затем происходит разделение клетки. При этом генетический материал равномерно распределяется между двумя идентичными в геноти-пическом плане клетками. Это свойство клеток является не только основой возникновения, развития организма, но и поддержания его целостности на протяжении всей жизни. Огромное значение деление клеток имеет и при регенерации поврежденных тканей.
Наиболее распространенной формой самовоспроизведения клетки является митоз, на морфологических проявлениях которого мы и остановимся несколько ниже. Первоначально необ-
Клетка
21
 ходимо охарактеризовать стадию, предшествующую митозу, а именно интерфазу (рис. 1.1.31, 1.1.32).
ходимо охарактеризовать стадию, предшествующую митозу, а именно интерфазу (рис. 1.1.31, 1.1.32).
D
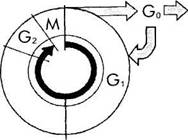
Рис. 1.1.31. Клеточный цикл:
| 14 |
 G,, G2 и G,, — периоды интерфазы; М — митоз; D — гибель клетки
G,, G2 и G,, — периоды интерфазы; М — митоз; D — гибель клетки
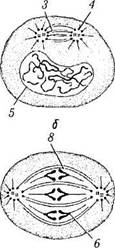
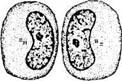
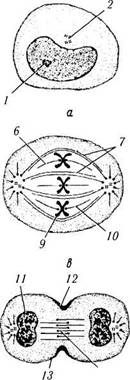
Рис. 1.1.32. Фазы митотического деления (по А. Хэм, Д.Кормак, 1982):
а — интерфаза (G2); 6 —профаза; в — метафаза; г — анафаза; д — телофаза; е — интерфаза (G,) (/ — ядрышко; 2 — центрио-ли; 3 —веретено деления; 4 —звезда; 5—ядерная оболочка; 6 —кинетохор; 7 — непрерывные микротрубочки; 8 — s-хромо-сома; 9 — d-хромосома; 10 — хромосомные микротрубочки; // — формирование ядра; 12 — борозда дробления; 13 — пучок актино-вых нитей; 14 — остаточное (срединное) тельце)
Интерфаза характеризуется наличием последовательных структурных и биохимических преобразований, подготавливающих клетку к митозу. Весьма важным в интерфазе является матричный синтез ДНК и удвоение хромосом — S-фаза. Промежуток времени между де-
лением и наступлением S-фазы называется фазой G, (постмитотическая или постсинтетическая фаза), а между S-фазой и митозом — фазой G2 (постсинтетическая или предмитотическая фаза). В течение фазы G, клетка диплоидная, в течение фазы S плоидность возрастает до четырех. Таким образом, в фазе G9 клетка уже тетраплоидная.
В интерфазе усиливаются биосинтетические процессы. Удваивается масса клетки, происходит деление центриолей. В течение предмитоти-ческой фазы (G2) обе материнские центриоли осуществляют сборку микротрубочек, усиливается формирование лизосом, делятся митохондрии и синтезируются новые белки, необходимые для осуществления митоза. К концу интерфазы хроматин конденсирован, ядрышко хорошо видно, ядерная оболочка не нарушена.
Наиболее важным и сложным процессом, происходящим в интерфазе, является удвоение набора хромосом. Суть удвоения состоит в том, что на цепочке ДНК синтезируется точно такая же параллельная цепочка. Этот процесс называется репликацией. Биологическая суть репликации сводится к тому, что при этом происходит передача генетической информации, хранящейся в родительской ДНК, путем точного ее воспроизведения в дочерней клетке. При этом каждая родительская цепь ДНК является матрицей для синтеза дочерней (матричный синтез ДНК). Процесс репликации довольно сложен.
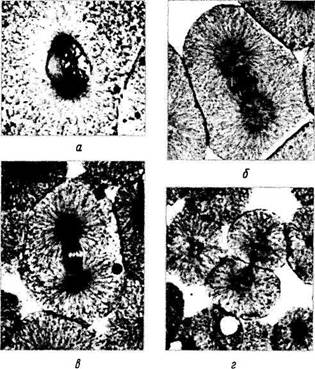
Рис. 1.1.33. Микроскопия фаз митоза на примере клеток бластулы (по Copenhaver, 1971):
а — поздняя профаза; б — метафаза; в — раняя анафаза; г — телофаза
22
Глава 1. КЛЕТКА И ТКАНИ
 Подробное описание этого процесса можно найти в многочисленных руководствах по генетике.
Подробное описание этого процесса можно найти в многочисленных руководствах по генетике.
В конце интерфазы клетка практически подготовлена к митотическому делению, которое в последующем и наступает. Морфологически различают 4 фазы митоза: профаза, метафаза, анафаза и телофаза (рис. 1.1.32, 1.1.33).
Профаза характеризуется тем, что в этот период происходят интенсивные изменения структуры ядра. На участках эухроматина прекращается транскрипция. Они покрываются белками и становятся неотличимыми от зерен гетерохроматина. Затем наступает спирализа-ция хромосом. При этом хромосома становится видимой в световом микроскопе. Вышеуказанный процесс сопровождается исчезновением ядрышка. Таким образом, в начале профазы в ядре образуется плотный клубок, который к концу фазы разрыхляется, и становятся видимыми хромосомы.
Именно в этой фазе центриоли расходятся к противоположным полюсам клетки, формируется веретено, состоящее из микротрубочек.
Метафаза (следует за профазой). В мета-фазе основные изменения происходят в цитоплазме клетки. Лизосомальные ферменты растворяют ядерную оболочку, и спирализованные хромосомы оказываются в цитоплазме. Комплекс Гольджи и эндоплазматический ретикулум распадаются на мелкие фрагменты.
На каждом центромере выявляется скопление специальных белков — кинетохор. Сборка микротрубочек на материнских центриолях продолжается. В результате этого процесса формируется биполярное митотическое веретено, состоящее из микротрубочек и ассоциированных с ними белков. Различают несколько видов микротрубочек. Часть микротрубочек расходится от центриоли во все стороны. Часть их образует астральную лучистость. Другая их часть направлена к экватору клетки — полярные микротрубочки. Кроме астральных и полярных микротрубочек от полюсов отходят ки-нетохорные микротрубочки, т. е. те, которые в области экватора прикрепляются к кинетохо-рам хромосом. В клетках человека каждый кинетохор связан с 20—40 микротрубочками.
Этап формирования веретена обозначают как прометафазу. В ходе собственно метафазы хромосомы перемещаются и располагаются в одной плоскости перпендикулярно к оси между полюсами. Образуется фигура, называемая материнской звездой. В результате упорядочения положения хромосом система микротрубочек также упорядочивается. Они теперь образуют веретено деления (митотическое веретено).
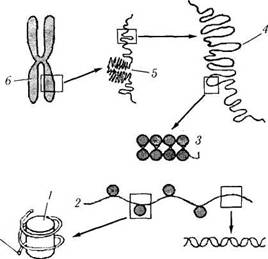
Важно отметить, что именно в метафазе определяют кариотип. У человека в норме ка-риотип характеризуется наличием 23 пар хромосом, приведенных на рис. 1.1.34. В метафазе изучают кариотип с диагностической целью при
| •1 mm |
U lift
Ну
| ДНК |
 fl/7 П
fl/7 П
| й | in | 2 | К II | и | ii 5& | ||
| X | 6 | 7 8 | 9 | 10 | И 12 | ||
| пл | Аи | Л6 | га | АЛ | |||
| 13 | 14 | 15 | 16 | 17 | 18 | 19 20 | |
| л | А А | ||||||
| Y | 21 | 22 |
ДНК
Рис. 1.1.34. Кариотип человека и уровни упаковки хроматина в ядре:
а — классификация пар хромосом человека; б — молекулярная организация хромосом (по В. Л. Быкову. 1999) двойная спираль ДНК образует нить диаметром 2 нм, которая намотана на блоки дисковндшш фирмы — нуклеосомы (/), входящие в состав нук-леосомнои пиiи (2) диаметром 11 нм Скрученная нуклеосомнля нить образует хроматиновую фибриллу (3) диаметром 30 нм, которая формирует петельные домены (4) диаметром 300 нм. Более и.ютно упакованные петельные до.мены образуют конденсированные участки хромосомы (5) диаметром 700 нм, являющими ч.н тью метафазной хромосомы (б) размером около 1400 нм
различных врожденных и наследуемых заболеваниях.
Анафаза. Анафаза довольно сложный в химическом отношении процесс. В начале анафазы наступает внезапное разделение центромеры d-хромосомы, в результате чего сестринские хроматиды становятся самостоятельными s-xpo-мосомами.
Микротрубочки начинают укорачиваться, в результате чего хроматиды подтягиваются к центриолям. Сами центриоли удаляются друг от друга в сторону полюсов клетки, в результате чего образуются две дочерние звезды. В конце анафазы плазматическая мембрана как бы инвагинируется перпендикулярно к продольной оси митотического веретена, образуя борозду. В этой области под плазмолеммой появляется сократительное кольцо, состоящее из
Клетка
23

|
| a U |
актин- и миозинсодержащих нитей. Завершает деление телофаза.
Телофаза. Как указано выше, к концу анафазы и началу телофазы в середине клетки образуется цитоплазматическая перетяжка, которая постепенно углубляется и, в конце концов, полностью разделяет клетку на две равные части, содержащие идентичный набор хромосом. После этого вновь появляется ядро, а хромосомы «распадаются» с образованием глыбок хроматина. Примерно в середине телофазы начинается образование нитчатой, а затем гранулярной частей нуклеонеммы. К концу телофазы полностью сформировано ядрышко. Из мембранных пузырьков происходит формирование аппарата Гольджи, эндоплазматической сети. На этом митотический цикл завершается, и клетка входит в интерфазу.
Вышеприведенные данные характеризуют основной тип клеточного деления — митоз. Но существуют и другие типы деления. Это эндомитоз. Морфологически при эндомитозе удвоение числа хромосом происходит внутри ядерной оболочки без ее разрушения и формирования веретена деления. При повторных эндомитозах число хромосом в ядре может значительно увеличиваться и развивается так называемая полиплоидия. Сопровождается этот процесс значительным увеличением объема ядра. Полиплоидия сопровождается значительным увеличением функциональной активности клеток. Полиплоидия характерна как в норме, так и при различных патологических состояниях эндотелия роговой оболочки.
Полиплоидия развивается и при митоти-ческом делении, при котором не происходит цитотомии. При последующем делении такой двуядерной клетки хромосомные наборы ядер объединяются в метафазе, приводя к образованию двух дочерних полиплоидных клеток. Наличие полиплоидных клеток (тетра-, окта- и т.д.) является нормальным состоянием ряда тканей организма человека.
Большое биологическое значение имеет еще один тип деления — мейоз, в результате которого формируются половые клетки. Основной смысл мейоза сводится к делению, при котором достигается уменьшение количества хромосом в клетке в два раза. Обсуждение этого типа деления выходит за рамки данной книги. Более подробные сведения можно получить в большом количестве руководств по цитологии.
В литературе описан еще один тип деления — амитоз. До сих пор обсуждаются вопросы возможности существования подобного типа деления. Считают, что при таком делении исчезает биологический смысл деления, т. е. возможность равного распределения генетического материала в двух вновь образованных клетках. Тем не менее морфологи, особенно патологи, довольно часто наблюдают прямое (амитотичес-кое) деление.
1.1.5. Межклеточные соединения
Межклеточное пространство. Между цито-плазматическими мембранами соседних клеток обнаруживается равномерное светлое пространство шириной 15 нм. Это пространство нередко расширяется или сужается как в норме, так и при патологических состояниях. Несмотря на наличие межклеточного пространства, клетки довольно сильно сцеплены между собой при помощи специализированных органоидов различного типа (рис. 1.1.35). На особенностях строения этих органоидов мы и остановимся ниже.
Рис. 1 1.35. Cxi'Mii гическое изображение межклеточных контактов различного типа (по tiogan et al., 1972):
ii.iuiiii.h- кл-динеиие; 2 —:ич мосома; 3 —ще.к-вой контакт I 'м.-I viz)); б — запмр.иощая зона [cmpt'-thu)
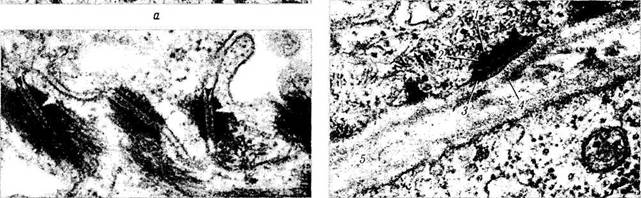
Десмосома (macula adhearens). Рядом расположенные клетки могут соединяться между собой при помощи локальных уплотнений — десмисим (рис. 1.1.36). Этот тип органоидов относится к адгезивным (контакты типа пятна слипания). При формировании подобного типа контакта цитоплазматические мембраны соседних клеток не сливаются, а как бы «прилипают» благодаря наличию межклеточного вещества. Особенно выражен подобный тип межклеточных контактов в эпителиальных тканях (эпидермис, эпителий роговицы, нейроэпите-лиальные структуры).
24
Глава 1. КЛЕТКА И ТКАНИ
|
|
|
границе со стромой) обнаруживаются не десмосомы, а полудесмосомы (рис. 1.1.37). По строению полудесмосома представляет собой как бы половинку десмосомы. Полудесмосома образована лишь одной пластинкой с входящими в нее тонофиламентами, которые прикрепляют клетку к базальной мембране. Подобные образования в большом количестве можно обнаружить в переднем и заднем эпителии роговой оболочки.
Рис. 1.1.36. Десмосомы:
а — светооптическая микроскопия плоскостного препарата однослойного плоского эпителия (между клетками видны десмосомы в виде мостиков); б —ультраструктурные особенности десмосом
Десмосомы настолько сильно связывают клетки между собой, что их можно разделить только механически. Даже при образовании эпителиальных пузырей на поверхности роговой оболочки (буллезная кератопатия), т.е. отслоения эпителиального пласта от подлежащей стромы, эпителиальный пласт долго сохраняет свою структуру именно благодаря десмосомам.
При ультраструктурном исследовании десмосомы представляют собой противостоящие дисковидные уплотнения мембран клеток (пластинка прикрепления). Диаметр этих пластинок около 0,5 мкм, а толщина 15 нм. Между ними определяется светлое межклеточное пространство (ширина 30 нм), выполненное «цементной» субстанцией умеренной электронноплот-ности в виде полоски. В межклеточном материале десмосомы определяются трансмембранные Са2+ связывающие адгезивные белки (десмо-коллины, десмоглеины), которые, взаимодействуя с белками пластинок прикрепления, связывают их в единую систему. В центре межклеточной щели видно линейное уплотнение (центральная или промежуточная линия). Радиаль-но от участков уплотнения распространяются внутрицитоплазматические промежуточные фи-ламенты, состоящие из особых белков, — дес-моплакины, плакоглобин и десмокальмин.
При контакте эпителиальной клетки с межклеточным материалом (базальная мембрана на
Рис. 1.1.37. Ультраструктурная организация полудесмосомы и базальной мембраны:
/ — полудесмосома; 2 — промежуточные филамеиты; 3 — плаз-молемма; 4 — якорные фибриллы; 5 — базальная мембрана
Промежуточное соединение, или опоясывающая десмосома (zonula adherens — поясок сцепления).
Подобного типа соединения обнаруживаются чаще всего на боковой поверхности эпителиальных клеток между областью расположения плотного соединения и десмосом. Это соединение охватывает клетку по периметру в виде пояска. В области промежуточного соединения обращенные к цитоплазме листки плазмолеммы утолщены и образуют пластинки прикрепления, которые содержат актин-связывающие белки альфа-актин, винкулин и плакоглобин. К этим пластинкам прикрепляются актиновые мико-филаменты, вплетающиеся также в терминальную сеть. Межклеточная щель расширена до 15—20 нм и заполнена умеренно электронно-плотным веществом, состоящим из адгезивного трансмембранного гликопротеина (Е-кадгерин) (рис. 1.1.38).
Плотное соединение (zonula occludens — поясок замыкания).
Этот тип контактов относится к так называемым плотным контактам. В контактах подобного рода цитоплазматические мембраны соседних клеток как бы сливаются. При этом образуется исключительно плотная стыковка клеток (рис. 1.1.39). Такие контакты наиболее часто встречаются в тканях, в которых необходимо полностью предотвратить проникновение метаболитов между клетками (эпителий кишеч-
Клетка
25

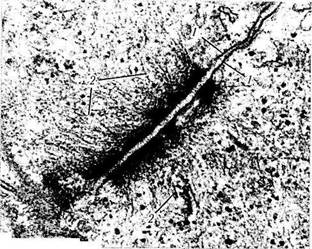
Рис. 1.1.38. Промежуточное соединение:
/ — цитоплазматические мембраны соседних клеток; 2 — микро-филаменты
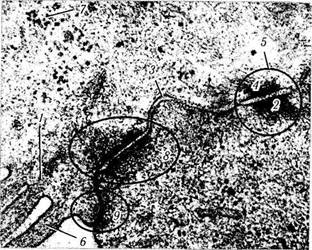
Рис. 1.1.39. Юнкциональный комплекс эпителиальных клеток:
/ — актиновые филаменты микроворсинок; 2 — цитоплазмати-ческая пластинка десмосомы; 3 — межклеточное пространство; 4 — промежуточные филаменты; 5 — десмосома (macula adherens); 6 — микроворсинка; 7 — полирибосомы; 8 — поясок сцепления (zonula adherens); 9 — плотное соединение (zonula occludens)
ника, эндотелий роговой оболочки). Как правило, соединения этого типа располагаются на апикальной поверхности клетки, опоясывая ее. Поясок замыкания представляет собой область частичного слияния наружных листков плазмолемм двух соседних клеток. Плотное соединение имеет вид пояска шириной 0,1 — 0,5 мкм, окружающего клетку по периметру и состоящего из анастомозирующих тяжей внут-римембранных частиц. Эти частицы образованы белком окклюдином. Каждая из них представляет собой область точечного слияния плазмолемм двух соседних клеток. Проницаемость плотных соединений тем ниже, чем выше число
тяжей этих частиц. Эти образования могут динамично перестраиваться и временно размыкаться.
В литературе нередко используется термин «юнкциональный комплекс». При этом авторы используют этот термин, описывая сложные межклеточные соединения, включающие одновременно типичные десмосомы, зону замыкания и ленту замыкания (fascia occludens) (рис. 1.1.39). Последний тип контакта отличается от зоны замыкания лишь тем, что контакт не полностью опоясывает клетку, а местами прерывается.
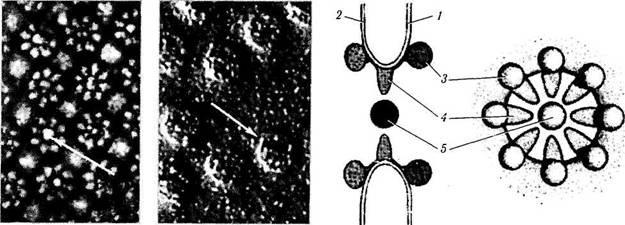
Щелевидное соединение (nexus). Щелевид-ное соединение характеризуется тем, что между цитоплазматическими мембранами соседних клеток видна щель шириной в 2 нм. При этом обе плазмолеммы соседних клеток соединены между собой коннексонами — полыми гексагональными белковыми структурами размерами около 9 нм, каждая из которых образована шестью белковыми субъединицами (рис. 1.1.40). Число коннексонов в щелевом соединении обычно исчисляется сотнями. Через эти образования осуществляется перенос метаболитов из одной клетки в другую. При этом молекулярный вес этих веществ не превышает 1500 Да (неорганические ионы, сахара, витамины, аминокислоты, нуклеотиды, АТФ и др.). Эти соединения обеспечивают между клетками ионное и метаболическое сопряжение. Близкое строение имеют синапсы.
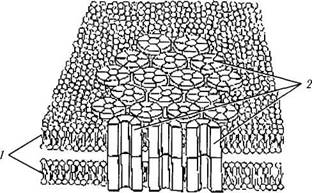
Рис. 1.1.40. Схематическое изображение щелевидного соединения:
/—цитоплазматические мембраны соседних клеток; 2 —кон-несоны
Щелевидные контакты играют важную роль в осуществлении функций клеток, особенно обладающих выраженной электрической активностью. По этой причине неудивительно, что большое количество подобных контактов обнаруживается между нейронами сетчатой оболочки (см. Сетчатка). Широко распространены они также в сердечной мышце и ткани центральной нервной системы.
26
Глава 1. КЛЕТКА И ТКАНИ
|
| Интердигитации (пальцевые соединения). Интердигитации представляют собой межклеточные соединения, образованные выпячиваниями цитоплазмы одних клеток, вдающимися в цитоплазму других (рис. 1.1.41). Между плаз-молеммами соседних клеток всегда остается |
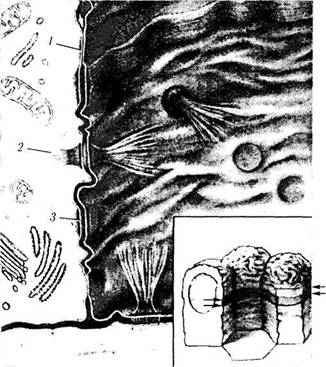
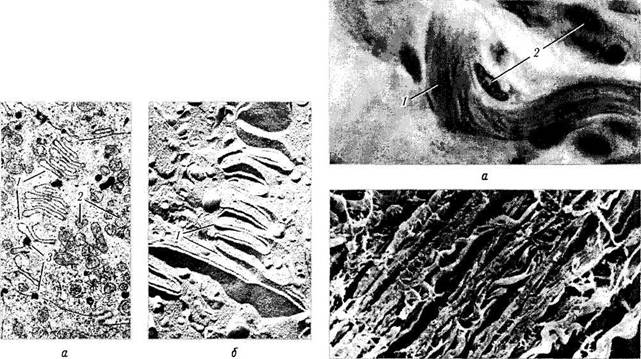
| Рис. 1.2.1. Коллагеновые волокна: а — плотная волокнистая ткань (виден пучок коллагеновых волокон (/) и фиброциты (2))\ б — ориентированные параллельно пучки коллагеновых волокон (сканирующая электронная микроскопия) |
Рис. 1.1.41. Интердигитации соседних эпителиальных клеток:
а — трансмиссионная электронная микроскопия (/ — интердигитации; 2 — митохондрии; 3 — десмосомы); б—метод замораживания-скалывания (сканирующая электронная микроскопия)
|
|
межклеточная щель 15—20 нм. За счет интердигитации увеличивается прочность межклеточного сцепления и увеличивается поверхность контакта между клетками.
1.2. МЕЖКЛЕТОЧНОЕ ВЕЩЕСТВО
Комплексы клеток вместе с межклеточным веществом образуют ткань. Межклеточное вещество состоит из волокон (коллагеновые, ретикулиновые, эластические), основного вещества. Некоторые авторы к нему также относят кристаллический материал и базальные мембраны.
Волокна
Коллагеновые волокна. Коллагеновое волокно является основным компонентом соединительной, хрящевой и костной тканей. При световой микроскопии видно, что коллагеновые волокна формируют различной толщины пучки, ориентированные в различных направлениях (рис. 1.2.1). При ультраструктурном исследовании отдельное коллагеновое волокно обладает поперечной исчерченностью (рис. 1.2.2).
Образование коллагеновых волокон происходит благодаря деятельности клеток соединительной ткани — фибробластов. Фибробласт рис. 1.2.2. Поперечная исчерченность коллагеновых во-синтезирует химические компоненты волокон, локон (трансмиссионная электронная микроскопия)
Межклеточное вещество
27
 а образование самого волокна происходит вне клетки путем сложных процессов самоорганизации. Самоорганизация волокна сводится к пространственной организации макромолекул. При этом образуются, в порядке усложнения, филаменты, фибриллы и волокна (рис. 1.2.3).
а образование самого волокна происходит вне клетки путем сложных процессов самоорганизации. Самоорганизация волокна сводится к пространственной организации макромолекул. При этом образуются, в порядке усложнения, филаменты, фибриллы и волокна (рис. 1.2.3).
|
|
D
I I I I I III
| 4 | (0,6D) | |
|
| ||
| * | ||
|
| ||
| 4.4Z) | ! |
|
|
300 нм
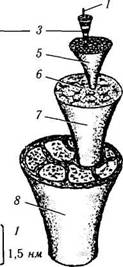
Рис. 1.2.3. Образование коллагеновой фибриллы и сборка фибрилл в коллагеновые структуры более высокого порядка (по В. Л. Быкову, 1999):
1 — молекула тропоколлагена; 2 — альфа-полипептидные цепи молекул тропоколлагена; 3 — коллагеновые фибриллы; 4 — зоны зазора; 5 — коллагеновые волокна; 6 — первичные коллагеновые пучки; 7 — вторичные коллагеновые пучки; 8 — третичный коллагеновый пучок (например, сухожилие)
Волокно состоит из пучков параллельно расположенных фибрилл, связанных цементирующим веществом. Толщина фибриллы порядка 5—10 нм. Каждая фибрилла, в свою очередь, состоит из нескольких протофибрилл, являющихся агрегатами волокнистого белка — коллагена. Основным компонентом волокон являются макромолекулы, состоящие из линейного, в виде цепочки, соединения различных аминокислот. Называется такая макромолекула тро-поколлагеном.
При этом коллагену свойствен особый аминокислотный состав. В коллагене обнаруживаются гликокол, пролин, оксипролин, глюта-миновая и аспарагиновая кислоты гидрокси-пролин и гидроксилизин. Две аминокислоты, гидроксипролин и гидроксилизин, составляют специфическую характеристику коллагеновой молекулы, т. к. они не встречаются в других веществах и структурах в значительной концентрации.
Размеры тропоколлагеновой молекулы следующие: диаметр—1,2—1,4 нм, длина — 280 нм.
Молекула коллагена содержит три полипептидные цепи, расположенные по спирали. В свою очередь каждая полипептидная цепь
содержит около 1000 аминокислотных остатков. Предполагают, что полипептиды образуют спираль с периодом 2,86 нм вокруг общей оси. Продольно ориентированные молекулы смещены в параллельных рядах на '/4 своей длины, с чем и связывают некоторые исследователи наличие поперечной исчерченности волокон, наблюдаемой как при электронной микроскопии, так и рентгеноструктурно. Так, рентгеновская дифракция выявляет, что нативный коллаген обладает периодичностью в 64 нм, аналогичной обнаруживаемой при электронной микроскопии.
Коллагеновая молекула химически связана с небольшим количеством Сахаров, гликопро-теинов.
К настоящему времени идентифицировано более 30 вариантов альфа-цепей коллагена, различных по химическому составу. Каждая из них кодируется различным геном. Хотя теоретически комбинации всех вариантов альфа-цепей могли дать более 1000 молекулярных форм коллагена, выявлено лишь 19 типов (обозначаются римскими цифрами). Эти коллагены различаются аминокислотным составом их альфа-цепей, порядком чередования в них аминокислот, молекулярной массой, распределением в тканях. Наибольшее значение имеют 5 первых типов коллагена. Коллагены I, II, III и V типов называются интерстициальными, или фибриллярными, так как они образуют фибриллы, которые входят в состав соединительной ткани. Коллагены различных типов широко представлены в тканях глаза. Строма роговой оболочки содержит всего пять разновидностей коллагена.
Важно упомянуть о некоторых особенностях использования терминологии при описании строения коллагеновых структур глаза. В тканях глаза термин «фибрилла» применяется для коллагена большого диаметра (роговица, уве-альный тракт, склера).
Термин «филамент» используется для нежных структур, а термин «волокно» используется для более грубых структур.
Основными функциями коллагеновых волокон являются следующие:
1. Обеспечение высоких механических
свойств соединительной ткани.
2. Определение архитектоники соединитель
ной ткани.
3. Обеспечение взаимодействия между клет
ками и межклеточным веществом.
4. Влияние на пролиферацию, дифференци-
ровку и функциональную активность многих
клеток.
Нарушение синтеза коллагеновых волокон приводит к существенному нарушению формирования соединительнотканных образований организма и развитию тяжелых заболеваний.
Ретикулярные волокна. Кроме коллагеновых волокон, в соединительной ткани (особенно рыхлой волокнистой неоформленной) обна-
28
Глава 1. КЛЕТКА И ТКАНИ

|
| - Ретикулярные волокна |
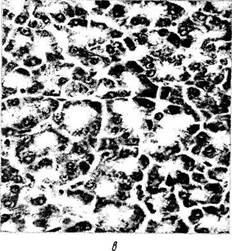
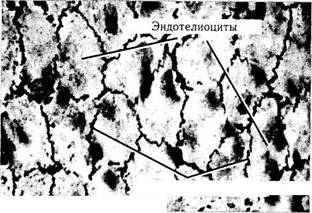 руживаются ретикулярные волокна (аргиро-фильные). Свое первое название эти волокна получили в результате того, что в тканях они формируют сеточку (рис. 1.2.4). Свое второе название волокна получили в связи с тем, что они выявляются после импрегнации ткани серебром.
руживаются ретикулярные волокна (аргиро-фильные). Свое первое название эти волокна получили в результате того, что в тканях они формируют сеточку (рис. 1.2.4). Свое второе название волокна получили в связи с тем, что они выявляются после импрегнации ткани серебром.
Ретикулярные волокна |
/
6
Рис. 1.2.4. Ретикулярные волокна, расположенные в межтрабекулярном пространстве паренхимы печени (а) и между клетками эндотелия кровеносного сосуда (б) (импрегнация серебром)
Ретикулярные волокна ориентированы в различных направлениях, формируя нежную сеть, окутывающую клетки. Являются они структурным компонентом базальных мембран (образуют их ретикулярную пластинку). Они также оплетают эпителиальные клетки, окружают кровеносные сосуды, нервные стволы.
Вырабатывать ретикулярные волокна способны, помимо фибробластов, жировые клетки,
гладкомышечные клетки, поперечно-полосатая мышца, мышечные клетки сердечной мышцы, шванновские клетки периферической нервной системы.
Ретикулярные волокна складываются из двух компонентов — цементирующего вещества (гликопротеины, протеогликаны) и микрофибрилл. При электронной микроскопии микрофибриллы представляют собой либо типичные коллагеновые фибриллы III типа (периодичность 64 нм), либо «атипичные» (периодичностью 22 нм). В стекловидном теле преобладают атипичные микрофибриллы.
В настоящее время различают собственно ретикулярные волокна и проколлагеновые. Собственно ретикулярные волокна являются конечной стадией развития волокна, а проколлагеновые — начальная стадия развития коллагено-вого волокна.
Основной функцией аргирофильных волокон является опорная функция. Они обнаруживаются во всех типах соединительной ткани, формируя поддерживающий клетки каркас (особенно в многоклеточных тканях типа лимфоид-ной, миелоидной).
Эластические волокна. Эластические волокна при микроскопическом исследовании хорошо выявляются только в тех случаях, когда они присутствуют в большом количестве. К подобным тканям относятся связки или внутренняя эластическая мембрана больших артерий. Немало этих волокон в увеальном тракте и склере глазного яблока.
Эластическая ткань идентифицируется при световой микроскопии после окраски специальными методами (орсеин, резорцин-фуксин). В отличие от коллагеновых волокон эластические не обладают поперечной исчерченностью (рис. 1.2.5).
В химическом отношении эластические волокна отличаются от коллагеновых более низким содержанием гидроксипролина и отсутствием гидроксилизина. Отмечается также высокое содержание валина. Волокна химически инертны. Электронномикроскопически эластическая ткань характеризуется наличием двух компонентов — фибриллярной части (микрофибриллы диаметром 11 нм) и гомогенной части (эластин). Гомогенная часть располагается в центре волокна и окружена микрофибриллами в виде трубочек.
Микрофибриллы и эластин отличаются друг от друга химическим составом. Эластин содержит аминокислоты десмозин и изодесмо-зин, обеспечивающие перекрестные полимерные связи, приводящие к формированию синцития. Микрофибриллы бедны этими аминокислотами. Они богаты полярными аминокислотами и углеводами.
Эластические волокна варьируют по толщине в пределах 0,2—10,0 мкм, ветвятся и анасто-мозируют друг с другом, формируя трехмер-
Межклеточное вещество
29

ЯЗШНШй |
| Поперечная сшивка молекул |
| * .'. |
| > |
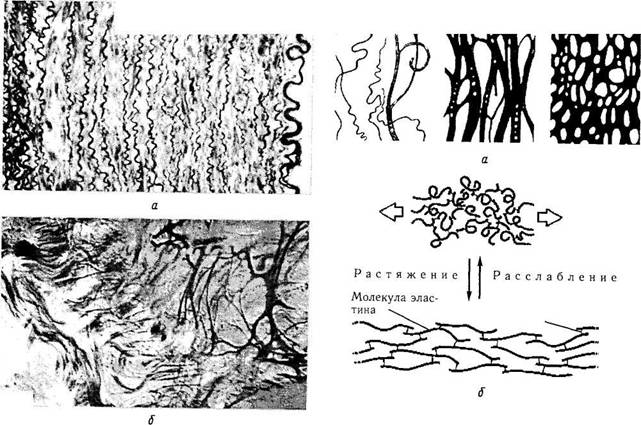
Рис. 1.2.5. Эластические волокна стенки аорты (а) и эластического хряща (б) (окрашивание орсеином)
ные сети (рис. 1.2.6). Особый тип связи между отдельными эластическими волокнами является структурной основой высокой способности эластической ткани к обратимой деформации.
Гистологи нередко используют термин «эластическая система». К этой системе, помимо эластических волокон, являющихся основным и наиболее зрелым элементом, относят также окситалановые и элауниновые волокна. Первые образованы микрофибриллами толщиной Ю—12 нм, сходными с теми, которые окружают центральный аморфный компонент эластического волокна. Вторые по строению занимают промежуточное положение между типичными эластическими и окситалановыми волокнами. В настоящее время считают, что окситалановые и элауниновые волокна являются предшественниками эластического волокна.
Помимо фибробласта эластическое волокно может образовываться в результате деятельности гладких миоцитов, хондробластов, хонд-роцитов.
В глазном яблоке преобладают эластопо-добные фибриллы, а именно так называемые фибриллы, состоящие из фибриллина. По этой причине мы несколько более подробно остановимся на этих фибриллах.
Фибриллин представляет собой гликопро-теид, являющийся основным компонентом рес-
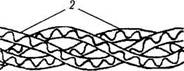
Рис. 1.2.6. Схематическое изображение организации эластической ткани:
а — различные типы организации эластических волокон (слева направо: эластические волокна, пучок эластических волокон, эластическая сеть); б—механизм обратимой деформации эластической ткани
ничного пояска хрусталика (связка Цинна). Фибриллин играет также важную роль в адаптационной способности соединительной ткани глаза.
Фибриллин полимеризуется с образованием микрофибрилл [19, 25, 28], которые складываются в параллельные пучки, образуя волокна ресничного пояска [38]. Богатые фибриллином микрофибриллы ресничного пояска морфологически идентичны микрофибриллам, которые обеспечивают эластические свойства и других тканей организма человека, таких как соединительная ткань кровеносных сосудов, легкого связок, дермы [22, 32]. Но в перечисленных тканях богатые фибриллином микрофибриллы являются основой для осаждения и правильной ориентации тропоэластина, предшественника эластина [13, 27]. Заключительным этапом этого процесса является образование эластических волокон. В глазном яблоке, особенно в области ресничного пояска, не содержится достаточно большого количества эластина [21].
Кроме ресничного пояска, в глазном яблоке фибриллин обнаружен также в соединительнотканных образованиях переднего отдела глаза, включая конъюнктиву, радужную оболочку глаза, строму ресничного тела, ресничные отростки, строму роговицы, базальную мембрану
30
Глава 1. КЛЕТКА И ТКАНИ

Гиалуроновая кислота Хондроитинсульфат, дераматансульфат Гепарансульфат, гепарин Кератансульфат |
эпителия роговицы, эндотелии шлеммова канала [35].
В заднем сегменте глаза фибриллин выявляется в строме склеры, решетчатой пластинке, мембране Бруха и сосудистой оболочке [35]. Выявляется он также в стекловидном теле [11, 34, 37].
Фибриллин существует в двух изоформах — фибриллин-1 и фибриллин-2. Контроль синтеза фибриллина-1 представлен в хромосоме 15q 15-21, а ген фибриллина-2 располагается в хромосоме 5q23-31 [20]. Фибриллин-1 и фибриллин-2 отличаются характером формирования микрофибрилл, и до сих пор непонятно, могут ли два этих типа гликопротеинов существовать в одной микрофибрилле или они образуют разные микрофибриллы [39, 40]. Предполагают, что фибриллин-2 играет основную роль в элас-тогенезе, а фибриллин-1 не участвует в элас-тогенезе, а обеспечивает эластические свойства тканей. Микрофибриллы ресничного пояска состоят исключительно из фибриллина-1 [21, 38].
Такое большое внимание фибриллину уделено нами по той причине, что аномалии синтеза этих микрофибрилл приводят к ряду заболеваний глаза. К таковым, в первую очередь, необходимо отнести эктопию хрусталика, синдром Марфана. При этом при эктопии хрусталика уменьшается количество волокон ресничного пояска, они растянуты и имеют различный диаметр [14, 23, 24]. Волокна легко разрушаются [19, 24].
Синдром Марфана, помимо наличия эктопии хрусталика, характеризуется развитием миопии, катаракты, открытоугольной глаукомы [17], косоглазия [16], плоской роговицы и гипоплазии ресничной мышцы и радужной оболочки [10, 12]. Выявляется также удлинение ресничных отростков [24, 26]. В последнее время показано также, что при псевдоэксфолиа-тивном синдроме, который иногда сопровождается вывихом хрусталика, эксфолиативный материал дает положительную реакцию при проведении иммуноморфологического выявления фибриллина [15, 29, 30].
Эти примеры последствий врожденных нарушений синтеза фибриллина довольно убедительно показывают большую роль этого глико-протеида в функционировании органа зрения.
Основное вещество
Основное вещество при гистологическом исследовании имеет вид студнеобразной массы, выполняющей межклеточные и межволоконные пространства соединительной ткани. В зависимости от типа соединительной ткани количество основного вещества различно. Наибольшее его количество в стекловидном теле.
Представлено основное вещество различными типами протеогликанов и структурных гликопротеинов.
Функциями протеогликанов соединительной ткани являются метаболическая (участие в транспорте метаболитов) и структурная (обеспечение структурной целостности волокнистого компонента). Структурная функция обеспечивается способностью протеогликанов взаимодействовать с молекулами коллагена, способствуя правильной укладке молекул тропоколлагена в фибриллах и фибрилл в волокнах. Протеоглика-ны обеспечивают также связь между поверхностью клеток и компонентами межклеточного вещества (фибронектином, ламинином, коллагеном). Протеогликаны обеспечивают также транспорт электролитов и воды благодаря способности связывать ее молекулы.
Протеогликаны состоят из пептидной цепи, связанной с гликозаминогликанами.
Гликозаминогликаны представляют собой неразветвленные отрицательно заряженные гидрофильные полисахаридные молекулы, образованные повторяющимися дисахаридными единицами. Основными гликозаминогликанами в организме человека являются гиалуроновая кислота, хондроитинсульфат, дерматансульфат, ге-парансульфат, гепарин, а также кератансуль-фат. В различных тканях и органах может преобладать один из типов гликозаминогликанов (табл. 1.2.1).
Таблица 1.2.1. Распределение гликозаминогликанов в организме человека
| Органы и ткани |
 Гликозаминогликаны
Гликозаминогликаны
Хрящ, синовиальная жидкость, кожа, пуповина, стекловидное тело, аорта
Хрящ, кость, кожа, кровеносные сосуды, сердце
Базальные мембраны, аорта, артерии легкого, легкое, печень, кожа, гранулы тучных клеток
Хрящ, роговица, межпозвонковый диск (студенистое ядро)
 Гликозаминогликаны, за исключением гиалу-роновой кислоты, связываются с белками, образуя протеогликаны.
Гликозаминогликаны, за исключением гиалу-роновой кислоты, связываются с белками, образуя протеогликаны.
Протеогликаны синтезируются в шероховатом эндоплазматическом ретикулуме, после чего выделяются в межклеточное пространство при помощи экзоцитоза. В межклеточном пространстве они объединяются в крупные проте-огликановые агрегаты. Протеогликаны разрушаются рядом лизосомальных ферментов клеток соединительной ткани.
Структурные гликопротеины представляют собой нефибриллярные белки, которые способствуют образованию базальных мембран, формированию фибрилл в межклеточном веществе. Эти вещества характеризуются разветвленной пептидной цепью, с которой связано
Межклеточное вещество
31
большое количество простых гексоз. К наиболее важным структурным гликопротеинам относятся фибронектин, ламинин и энтактин/ни- доген. Фибронектин синтезируется фиброблас-тами и другими клетками мезенхимного происхождения, а также эпителиальными клетками. Он обеспечивает организацию компонентов межклеточного вещества. Ламинин — гликопро-теин, входящий в состав базальных мембран. Энтактин/нидоген связывается с коллагеном IV типа и ламинином, входя в состав плотной пластинки базальной мембраны.
1.2.3. Кристаллические материалы
К наиболее распространенным кристаллическим материалам тканей относятся соли кальция. В норме в глазном яблоке кальцифи-каты никогда не обнаруживаются. Их отложение отмечается лишь при старении и ряде патологических состояний (ретинобластома, посттравматическая атрофия глаза).
1.3. БАЗАЛЬНЫЕ МЕМБРАНЫ
Базальной мембраной называют электронно-плотную структуру, связанную с базальной плазматической мембраной эпителиальной клетки, но лежащую вне клетки (рис. 1.3.1, 1.3.2). Базальная мембрана может быть очень тонкой,
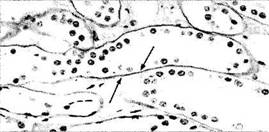
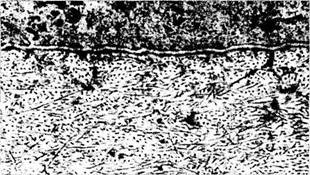
Рис. 1.3.1. Светооптическое (а) и ультраструктурное (б) строение базальной мембраны:
а — базальная мембрана (стрелка) эпителия почечных канальцев; б — ультраструктура базальной мембраны переднего эпителия роговой оболочки (стрелкой указаны якорные фибриллы)
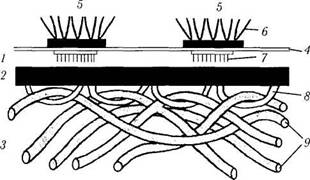
Рис. 1.3.2. Схематическое изображение строения базальной мембраны и полудесмосомы (по В. Л. Быкову, 1999):
1 — светлая пластинка; 2 — плотная пластинка; 3 — ретикулярная пластинка; 4 — плазмолемма; 5 — полудесмосома; 6 — промежуточные филаменты; 7 —якорные филаменты; 8 —якорные фибриллы; 9 — коллагеновые фибриллы
в такой степени, что различать ее при световой микроскопии не представляется возможным. Встречаются и толстые мембраны. Толстые базальные мембраны получили название «стекловидные мембраны». Существуют и базальные мембраны, видимые невооруженным глазом (капсула хрусталика).
Толстые базальные мембраны глаза представляют собой множество переплетающихся тонких базальных мембран, складывающихся в сложную многослойную структуру. Многослойные базальные мембраны могут быть составлены из толстых пластинок (периферия рогович-ного эпителия) или из тонких пластин (внутренняя пограничная мембрана ресничного эпителия).
Некоторые базальные мембраны (капсула хрусталика) обладают четкой волокнистой структурой.
Базальные мембраны прозрачны, обладают эластическими свойствами, способны к сокращению и сворачиваются при их разрушении (сворачивание десцеметовой оболочки после проникающего ранения роговицы).
Свободные поверхности толстых стекловидно подобных базальных мембран гладкие. По этой причине они интенсивно отражают свет. Этим объясняется блестящая поверхность десцеметовой оболочки, капсулы хрусталика, пограничной мембраны сетчатки.
Ультраструктурные исследования выявили, что базальные мембраны имеют довольно сложное строение. В них выделяют три слоя.
Первый слой — светлая пластинка (lamina lucida). Этот слой имеет толщину 30—50 нм и прилежит к плазмолемме базальной поверхности эпителиоцитов. От полудесмосом эпите-лиоцитов в глубь этой пластинки направляются тонкие якорные филаменты. Светлая пластинка содержит гликопротеины (в том числе сульфа-тированный гликопротеин ламинин) и антиген
32
Глава 1. КЛЕТКА И ТКАНИ
 пузырчатки (способствующие прикреплению базальной части эпителиоцитов), а также про-теогликаны (гепарансульфат).
пузырчатки (способствующие прикреплению базальной части эпителиоцитов), а также про-теогликаны (гепарансульфат).
Второй слой — плотная пластинка (lamina densa). Этот слой имеет толщину 50—60 нм и состоит из гранулярного и фибриллярного материала. Этот слой обращен в сторону эпителиальной ткани. В эту пластинку вплетаются якорные фибриллы, имеющие вид петель (образованы коллагеном VII типа), в которые продеты коллагеновые фибриллы подлежащей соединительной ткани. Плотная пластинка содержит коллаген IV типа, энтактин, гепарансульфат, коллаген V типа и адгезивный гликопротеин фибронектин.
Третья — ретикулярная — пластинка (lami na reticularis) состоит из коллагеновых фибрилл соединительной ткани, связанных с якорными фибриллами. В ее состав входят фибриллы, образованные коллагенами I и III типов. Хотя, по мнению некоторых авторов, эту пластинку не следует относить к собственно базальной мембране, именно она образует основную массу той структуры, которая выявляется ШИК-реакцией или окраской солями серебра.
Функциями базальной мембраны являются поддержание нормальной архитектоники, дифференциации и поляризации эпителия; обеспечение плотной связи эпителиоцитов с подлежащей соединительной тканью; избирательная фильтрация питательных веществ, обеспечение и регуляция роста эпителия по подлежащей соединительной ткани при его развитии и репа-ративной регенерации.
Нарушение строения и функции базальной мембраны приводит к развитию ряда заболеваний органов, включая глазное яблоко (диабетическая микроангиопатия).
1.4. ТКАНИ
Клетки и межклеточный материал образуют ткани. Ткань — это исторически сложившаяся система клеток и внеклеточных структур, обладающая общностью строения и специализированная на выполнение определенных функций [6]. Различают ряд типов тканей. Это эпителиальная ткань, кровь, соединительная ткань, мышечная и нервная ткани. Изучая микроскопически глаз, придаточный аппарат глаза и глазницу, можно встретиться со всеми перечисленными типами тканей. По этой причине имеет смысл кратко охарактеризовать особенности строения различных типов тканей.
Эпителиальная ткань
Основной особенностью эпителиальной ткани является ее расположение на границе с внешней или внутренней окружающей средой (покрывающая функция).
В структурном отношении эпителиальные ткани характеризуются скоплением клеток, плотно соединенных между собой межклеточными контактами.
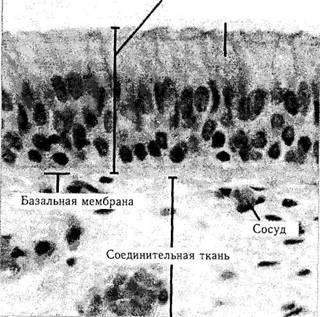
Слой эпителиальных клеток располагается на базальной мембране, под которой лежит соединительная ткань, богатая кровеносными сосудами.
В первую очередь эпителиальная ткань обеспечивает разграничительную, барьерную функцию. Эпителий образует барьеры между внутренней средой организма и внешней. Немногими исключениями из общего правила служат эпителии, разграничивающие две области внутренней среды, например выстилающие полости тела (мезотелий, эндотелий роговой оболочки) или сосуды (эндотелий). Производными основной разграничительной функции являются другие функции. Это защитная, транспортная (перенос метаболитов и др.), всасывающая, секреторная, экскреторная (удаление из организма продуктов обмена), сенсорная (восприятие механических, сенсорных, химических сигналов).
Эпителиальные ткани происходят из различных зародышевых листков — эктодермы (эпидермис, эпителиальная выстилка дыхательной системы, передний эпителий роговой оболочки и др.), мезенхимы (выстилка кровеносных сосудов и др.), энтодермы (эпителий кишечника и др.), нейроэпителия (эпендимная глия, выстилка мозговых оболочек и др.). Необходимо отметить, что в офтальмологии термин «эпителий» распространяется и на такие производные нейроэпителия, как пигментный эпителий сетчатки, ресничного тела, радужной оболочки.
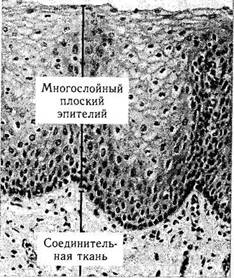
В структурном отношении эпителиальные ткани подразделяются на однослойный и многослойный эпителий (рис. 1.4.1 —1.4.3). Однослойные эпителии представляют собой один слой клеток, а многослойные состоят из большого количества плотно сцепленных между собой клеточных слоев.
Однослойные эпителии, в свою очередь, подразделяются на плоский (эндотелий роговицы), кубический (центральные зоны эпителия капсулы хрусталика), цилиндрический. То есть учитывается форма клеток.
Многослойные эпителии разделяются на многослойный плоский ороговевающий (эпителий кожи век), многослойный плоский неорого-вевающий эпителий (передний эпителий роговой оболочки, эпителий конъюнктивы). В последних случаях учитывается как форма поверхностно лежащего слоя эпителиальных клеток, так и появление специализированной функции, т. е. синтез кератогиалина.
Существует еще ряд реже встречающихся морфологических типов эпителиальной ткани, являющихся вариантами строения вышеперечисленных двух типов (многорядный, многослойный цилиндрический, переходный и др.).
Ткани
33
м
It*
I







 Рис. 1.4.1. Схематическое изображение различных морфологических типов эпителиальной ткани:
Рис. 1.4.1. Схематическое изображение различных морфологических типов эпителиальной ткани:
/ — однослойный эпителий; 2 — однослойный кубический;,?—однослойный (однорядный) призматический; 4 — однослойный много
рядный призматический; 5 — многослойный плоский неороговевающий; 6 — многослойный плоский ороговевающий; 7 многослой
ный кубический; 8 — переходный
|
|
|
|
| *. |
 6
6
 Эпителий
Эпителий
Бокаловидная клетка
Рис. 1.4.2. Микроскопические особенности некоторых типов эпителиальной ткани:
а — многослойный плоский эпителий; б — призматический эпителий; в — однослойный цилиндрический эпителий альвеолярной
железы; г —многослойный призматический эпителий
34
Глава 1. КЛЕТКА И ТКАНИ

|
|
|
|
|
Дата добавления: 2018-10-18; Мы поможем в написании ваших работ!; просмотров: 344 | Нарушение авторских прав
Лучшие изречения:
 | 4185 -
| 4185 -