




Вышеперечисленные структуры цитоплазмы объединены в единую группу по одной причине — элементарной составной их единицей является микротрубочка. Выявляются микротрубочки во всех типах клеток, за исключением бактерий. В структурах глаза, в частности в сетчатой оболочке, микротрубочки обнаруживаются в большом количестве. Основная функция микротрубочек — опорная, т. е. обеспечение определенной формы клетки и ее жесткости. По этой причине микротрубочки относят к структурам цитоскелета. Кроме того, они участвуют во внутриклеточном переносе метаболитов.
|
|

| 5 нм |
Микротрубочки имеют диаметр порядка 20—30 нм. Длина их различная. На поперечном срезе они имеют вид кольца (рис. 1.1.13). Каждая микротрубочка состоит из 13 протофила-ментов, расположенных вдоль длинной оси трубочки и скрученных по спирали одна над другой. Протофиламенты состоят из особого белка — тубулина. Сборка микротрубочки происходит из димеров тубулина (рис. 1.1.14). Синтез тубулинов происходит на мембранах грануляр-
Рис. 1.1.13. Схема структурной организации микротрубочки (по В. Л. Быкову, 1999):
а — мономеры тубулина, образующие протофиламенты; б — микротрубочка; s — пучок микротрубочек
Таблица 1.1.2. Морфо-функциональная организация митохондрий



 Структуры митохондрий
Структуры митохондрий
 Наружная мембрана Межмембранное пространство
Наружная мембрана Межмембранное пространство
Внутренняя мембрана
Субмитохондриальные частицы Матрикс
Состав
Около 20% всего белка митохондрий. Ферменты липидного обмена
Ферменты, использующие АТФ для фосфорилирования других нуклеоти-дов
Ферменты дыхательной цепи, цито-хромы. Сукцинатдегидрогеназа. Трансбелки.
АТФ-синтетаза
Ферменты (кроме сукцинатдегидро-геназы). ДНК, РНК, рибосомы, ферменты, участвующие в экспрессии генома митохондрий
Функция
Транспорт. Превращение липидов в промежуточные метаболиты
Создание электрохимического протонного градиента. Перенос метаболитов в матрикс и из него
Синтез и гидролиз АТФ
Цикл лимонной кислоты, превращение пирувата, аминокислот и жирных кислот в ацетил-коэнзим А. Репликация, транскрипция, трансляция
Клетка
11

|

мирующих сети. Микротрубочки могут образовывать пучки, в которых они связаны тонкими поперечными мостиками (в отростках нейронов, в составе митотического веретена и др.). Нередко микротрубочки частично сливаются, формируя пары (в аксонеме ресничек и жгутиков) или триплеты (в базальном тельце и цент-риоли).
| 12 |
 Микротрубочки являются составной частью и другого органоида—реснички (рис. 1.1.14, 1.1.15). Реснички располагаются на апикальной поверхности многих клеток, в основном эпителиальных, выстилающих влажные поверхности тканей. В клетке может быть одна или несколько сотен ресничек. Обычно ресничка имеет длину порядка 15 мкм, а диаметр — 0,2 мкм. В основании реснички располагается электрон-ноплотное образование, называемое базальным тельцем. Базальное тельце цилиндрическое и состоит из девяти пучков параллельных друг другу микротрубочек, по три в каждом пучке. Такой пучок, состоящий из трех микротрубочек, называется триплетом. Девять триплетов удерживаются фибриллярным материалом, образуя стенку цилиндра.
Микротрубочки являются составной частью и другого органоида—реснички (рис. 1.1.14, 1.1.15). Реснички располагаются на апикальной поверхности многих клеток, в основном эпителиальных, выстилающих влажные поверхности тканей. В клетке может быть одна или несколько сотен ресничек. Обычно ресничка имеет длину порядка 15 мкм, а диаметр — 0,2 мкм. В основании реснички располагается электрон-ноплотное образование, называемое базальным тельцем. Базальное тельце цилиндрическое и состоит из девяти пучков параллельных друг другу микротрубочек, по три в каждом пучке. Такой пучок, состоящий из трех микротрубочек, называется триплетом. Девять триплетов удерживаются фибриллярным материалом, образуя стенку цилиндра.
6 2
| ной эндоплазматической сети, а сборка в спирали — в клеточном центре. При этом поддерживается постоянное равновесие между сформированной микротрубочкой и растворенными в цитоплазме димеров тубулина, способных к самосборке. Эта закономерность не распространяется на постоянные органоиды клеток, состоящие из микротрубочек, — реснички, цент-риоли, базальные тельца. Нарушают процесс самосборки некоторые вещества, в частности колхицин и винбластин. Микротрубочки являются структурным компонентом веретена при делении клетки. Микротрубочки формируют в цитоплазме различные структурные системы. Они могут быть распределены в виде отдельных элементов, разбросанных по всей цитоплазме и фор- |
Рис. 1.1.14. Ультраструктурные особенности ресничек: а — продольный срез; б — поперечный срез
Рис. 1.1.15. Схематическое изображение организации реснички (по В. J1. Быкову, 1999):
а —продольный срез; б — поперечный срез (/ — базальное тельце; 2 — центр организации микротрубочек; 3 — базальный корешок; 4 — плазмолемма; 5 — микротрубочка А; 6 — микротрубочка В; 7 — периферические микротрубочки; 8 — центральные микротрубочки; 9 — центральная оболочка; 10 — динеино-вые ручки; // — радиальные спицы; 12 — нексиновые мостики)
Базальное тельце является организатором реснички. После образования базального тельца оно мигрирует к апикальной поверхности клетки. Из дистального конца базального тельца растут микротрубочки, составляющие стержень реснички (аксонема). Этот стержень, окруженный цитоплазматической мембраной, и вы-
12
Глава 1. КЛЕТКА И ТКАНИ
 стоит над поверхностью клетки. В центре аксо-немы образуются две одиночные микротрубочки, называемые центральной парой, или синг- летами.
стоит над поверхностью клетки. В центре аксо-немы образуются две одиночные микротрубочки, называемые центральной парой, или синг- летами.
Образованные реснички синхронно совершают движения, способствуя продвижению по эпителиальной поверхности слизистой оболочки секрета. Реснички эндотелия роговой оболочки обеспечивают перемещение камерной влаги в определенном направлении и с определенной скоростью, что имеет немаловажное значение в метаболизме структур глаза. Напоминающая ресничку структура с базальным тельцем обнаруживается и в фоторецепторных клетках.
Следующей структурой, состоящей из микротрубочек, является клеточный центр, образованный двумя полыми цилиндрическими структурами. Длина клеточного центра равна 0,3—0,5 мкм, а диаметр — 0,15—0,2 мкм. Каждая из этих структур называется центриолью. Располагаются они вблизи друг друга во взаимно перпендикулярных плоскостях недалеко от аппарата Гольджи (рис. 1.1.16). Часть цитоплазмы, где они лежат, называется центросомой.

и
Рис. 1.1.16. Клеточный центр и структурная организация центриоли:
а —ультраструктурные особенности клеточного центра (/— центриоль; 2 — комплекс Гольджи; 3 — десмосома; 4 — митохондрия; 5 — мембрана митохондрии; 6 — микротрубочки; 7 — шероховатый эндоплазматический ретикулум; 8 — гладкий эндо-плазматический ретикулум; 9 — межклеточное пространство) б — схема организации клеточного центра и центриоли (/ — центриоли; 2 — триплеты микротрубочек; 3 — микротрубочки; 4 — сателлиты)
Ультраструктурная их организация практически неотличима от строения базального тельца. Каждая центриоль состоит из 9 триплетов частично слившихся микротрубочек, связанных поперечными мостиками. Каждый триплет связан со сферическими тельцами (сателлитами). Расходящиеся от них микротрубочки образуют центросферу.
В неделящейся клетке выявляется одна пара центриолей (диплосома). Перед делением (S-фа-за) происходит дупликация центриолей пары, причем под прямым углом к каждой зрелой центриоли формируется новая (дочерняя), незрелая процентриоль. Пары центриолей затем расходятся к полюсам клетки. Во время митоза они служат центрами образования микротрубочек ахроматического веретена деления. Таким образом, основной функцией центриолей является участие в митотическом делении клетки.
Филаменты. В цитоплазме большинства клеток обнаруживается множество волокнистых структур (филаментов) (рис. 1.1.17, 1.1.18). Различают три типа филаментов (микрофила-менты, миозиновые филаменты и промежуточные филаменты).
Первый тип филаментов — это так называемые микрофиламенты. Диаметр их 5—б нм. В основном они состоят из белка актина. С актином связываются еще два типа белка, а именно тропомиозин и миозин. В результате этого процесса формируется актино-миозино-вый комплекс. При этом актин и миозин становятся способными смещаться в этом комплексе продольно относительно друг друга. Если концы комплекса скреплены с какими-либо другими внутриклеточными структурами, последние сближаются. Этот процесс лежит в основе перемещения внутри цитоплазмы органоидов, транспортных пузырьков и других структур. На этом основано и мышечное сокращение.
Микрофиламентов особенно много в поверхностных областях цитоплазмы (поверхностный комплекс). Тем самым они способствуют поступлению веществ в цитоплазму (пиноцитоз), обладая возможностью изменять конфигурацию плазмолеммы.
Актиновые филаменты прикрепляются к трансмембранным белкам в особых участках плазмолеммы, называемых адгезионными соединениями, или фокальными контактами, которые связывают клетки друг с другом или клетки с компонентами межклеточного вещества.
Второй тип филаментов называют миозино- выми филаментами, поскольку они состоят из белка миозина. Этот тип филаментов тесно связан с актиновыми микрофиламентами в мышечных клетках. Эти филаменты толще (диаметр равняется 10 нм).
Название третьего типа филаментов — про межуточные. Их диаметр колеблется от 7 до 10 нм. Промежуточные филаменты наиболее часто встречаются в нервных и глиальных клет-
|
|

|
|

|
|

Клетка
13

 25 мкм
25 мкм

25 нм
Рис. 1.1.17. Особенности распределения структурных элементов цитоскелета (верхняя часть рисунка) и их
молекулярная организация (нижняя часть рисунка):
а — промежуточные филаменты; б — микротрубочки; в — актиновые филаменты
 |
Необходимо отметить и то, что, несмотря на сходное строение, промежуточные филаменты отличаются в клетках различных тканей своим химическим составом (табл. 1.1.З.). Таблица 1.1.3. Распределение промежуточных фи ламентов различных классов в клетках и тканях человека |
| Химические особенности белков промежуточных филаментов легко демонстрируются иммуноцитохимическими методами, поскольку каждый белок является антигеном. Идентификация классов промежуточных филаментов имеет большое диагностическое значение при установлении тканевой принадлежности опухоли. Наибольшее диагностическое значение |

Рис. 1.1.18. Продольный срез отростка глиальной клетки сетчатки. Ультраструктурные особенности внутри-цитоплазматических филаментов
ках (в частности, в сетчатой оболочке, зрительном нерве). Эти филаменты в клетке образуют трехмерные сети. Входят они также в состав десмосом и полудесмосом эпителиальных клеток. К основным функциям промежуточных филаментов в настоящее время относят опорную функцию, обеспечение равномерного распределения сил деформации между клетками ткани (препятствует повреждению отдельных клеток), участие в образовании рогового вещества в эпителии кожи, поддержание формы отростков нервных клеток и фиксация трансмембранных белков. Кроме того, эти филаменты обеспечивают удержание миофибрилл в мышечной ткани и прикрепление их к плазмо-лемме (обеспечение сократительной функции мышц).
Классы промежуточных филаментов
 (Цито-)кератиновые (тонофиламенты)
(Цито-)кератиновые (тонофиламенты)
Десминовые
Виментиновые
Нейрофиламенты
Глиальные (содержат глиальный фибриллярный кислый белок)
Ламины (образуют ка-риоскелет)
Типы клеток и тканей
Эпителиальные
Мышечные ткани — гладкие (кроме миоцитов сосудов) и поперечнополосатые
Различные клетки мезенхим-ного происхождения: фибро-бласты, макрофаги, остеобласты, хондробласты, эндотелий и гладкие миоциты сосудов
Нейроны
Глиальные клетки (астроциты, олигодендроглиоциты)
Все типы клеток
14
Глава I. КЛЕТКА И ТКАНИ

 имеет выявление цитокератинов, десмина и глиального фибриллярного кислого белка, которые служат маркерами опухолей эпителиального, мышечного и глиального происхождения. Менее отчетливые результаты дает обнаружение виментина.
имеет выявление цитокератинов, десмина и глиального фибриллярного кислого белка, которые служат маркерами опухолей эпителиального, мышечного и глиального происхождения. Менее отчетливые результаты дает обнаружение виментина.
Микроворсинки. Некоторые (эпителиальные) клетки на своей апикальной поверхности содержат многочисленные цитоплазматические выросты, значительно увеличивающие площадь контакта содержимого цитоплазмы с окружающей средой, что способствуют увеличению интенсивности всасывания питательных веществ. Называются эти органоиды микроворсинками (рис. 1.1.5, 1.1.19). Естественно, что чаще подобные образования обнаруживаются в эпителиальных клетках, особенно клетках слизистой желудочно-кишечного тракта. Пигментный эпителий сетчатой оболочки также обладает подобными образованиями, распространяющимися между наружными члениками палочек и колбочек фоторецепторных клеток сетчатки. Многочисленны они и на апикальной поверхности эндотелиальных клеток роговой оболочки.
Микроворсинки имеют диаметр порядка 0,1 мкм. Длина их может быть самой различной.
В центральной части микроворсинки, представляющей собой выпячивание цитоплазмы, располагается порядка 40 микрофиламентов (диаметр 6 нм). В апикальной части микроворсинки пучок микрофиламентов закреплен в аморфном веществе. Его жесткость обуслов-
Рис. 1.1.19. Схема ультраструктурной организации микроворсинки:
/ — актиновые микрофиламенты; 2 — аморфное вещество; 3 — фимбрин и виллин (белки, образующие поперечные сшивки в пучке актиновых микрофиламентов); 4 — молекулы минимиозина (прикрепляющие пучок актиновых микрофиламентов к плазмолемме микроворсинки); 5 — терминальная сеть актиновых микрофиламентов; 6 — спектриновые мостики (прикрепляют терминальную сеть к плазмолемме); 7 — миозиновые филаменты; 8 — промежуточные филаменты; 9 — гликокаликс
лена наличием поперечных сшивок из белков фимбрина и виллина. Изнутри пучок прикреплен к плазмолемме микроворсинки белковыми мостиками из минимиозина.
Непосредственно под ворсинкой в цитоплазме обнаруживается скопление миозиновых фи-ламентов. Предполагается, что взаимодействие этих двух типов филаментов способствует изменению конфигурации микроворсинок, что еще более усиливает всасывательную функцию клеток.
Стереоцилии представляют собой видоизмененные длинные микроворсинки.
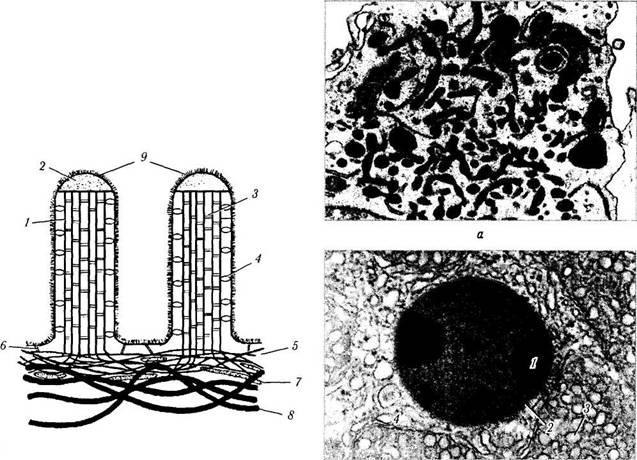
Лизосомы (цитосомы). Уже давно в большинстве клеток были обнаружены мембранные органоиды в виде «темных телец» различного размера (0,4—0,5 мкм) (рис. 1.1.20). Основным отличием этих образований было то, что они содержали целый набор (около 50) гидролаз. Поскольку гидролазы осуществляют лизис, эти органоиды и были названы «лизосомы» (переваривающие тельца).
Рис. 1.1.20. Лизосомы:
а — высокая степень насыщения цитоплазмы лизосомами, вблизи которых видны бобовидной формы митохондрии; б — ультраструктурные особенности вторичной лизосомы (/ — электронно-плотные включения в лизосому; 2 — мембрана лизосомы; 3 —митохондрии; 4 — гладкий эндоплазматический ретикулум)
Клетка
15

|

| Пиноцитоз-ные пузырь- Мультивезику- / ки Аутофаго- |
| Фагоцитоз |
| Первичная лизосома |
| ш Фагоцитоз |
Комплекс Гольджи
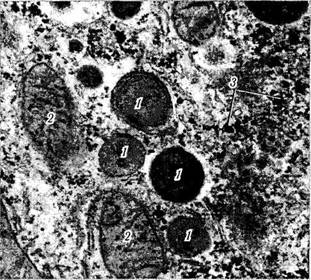
 0,25 мкм) присутствуют лишь в некоторых тканях (почки, печень). В них обнаруживается кристалловидная сердцевина, в которой находятся ферменты в концентрированной форме.
0,25 мкм) присутствуют лишь в некоторых тканях (почки, печень). В них обнаруживается кристалловидная сердцевина, в которой находятся ферменты в концентрированной форме.





