




Между внутренней и внешней стороной клетки всегда есть разность электрических потенциалов. Эта разность в состоянии физиологического покоя клетки называется потенциалом покоя.
Установлено, что возникновение потенциала покоя обусловлено в основном наличием концентрационного градиента ионов калия. Эта точка зрения базируется на том, что ионы калия внутри клетки находятся в свободном состоянии, т.е. не связаны с другими ионами и молекулами и могут свободно диффундировать. Ионы калия диффундируют по концентрационному градиенту через клеточную мембрану и остаются на ее внутренней стороне. Т.к. ионы калия имеют «+» заряд, а анионы «-» заряд, то внешняя поверхность мембраны заряжается положительно, а внутренняя отрицательно.

Понятно, что диффузия продолжается только до тех пор, пока не установится равновесие между силами возникающего электрического поля и силами диффузии.
Мембрана в состоянии покоя проницаема не только для ионов калия, но и в небольшой степени для ионов натрия и хлора. Мембранный потенциал клеток представляет собой результирующую электродвижущих сил, генерируемых всеми этими каналами диффузии. Возникающий при этом потенциал описывается уравнением Гольдмана – Ходжкина – Катца:

где  – коэффициенты проницаемости мембраны для ионов
– коэффициенты проницаемости мембраны для ионов  .
.
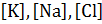 - концентрация ионов внутри (
- концентрация ионов внутри (  ) и вне (
) и вне (  ) клетки.
) клетки.
Таким образом, величина потенциала покоя клетки определяется двумя основными факторами:
1. соотношением концентраций проникающих через покоящуюся мембрану катионов и анионов;
2. соотношением проницаемостей мембраны для этих ионов.
Состояние покоя характеризуется следующим отношением ионных проницаемостей:
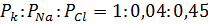
Уравнение Гольдмана переходит в уравнение Нернста, если мембрана проницаема для одного вида ионов. Следует отметить, что наличие обменной диффузии не влияет на мембранный потенциал.
Наиболее распространенный метод измерения биопотенциала покоя – микроэлектродный. При этом микроэлектроды вводятся: первый непосредственно в цитоплазму клетки, второй – во внеклеточную жидкость. Установлено, что цитоплазма в состоянии покоя клеток всегда имеет отрицательный потенциал по отношению к потенциалу межклеточной жидкости и колеблется в пределах от -100 милливольт до -50 милливольт.
6.3. Потенциал действия. Потенциал действия аксона нервной клетки.Уравнение Ходжкина – Хаксли.
Общее изменение разности потенциалов между клеткой и средой, происходящее при возбуждении клетки, называется потенциалом действия. В его основе лежат изменения ионной проницаемости мембраны. Потенциалы действия обеспечивают проведение возбуждения по нервным волокнам и инициируют процессы сокращения мышечных и секреции железистых клеток.
Представим график потенциала действия аксона нервной клетки. Потенциал как функция времени показан в фиксированной точке аксона.

φ 1= -70 мВ потенциал покоя
φ 2= +40 мВ мембранный потенциал при возбуждении
φ 3 = / φ 1/+ φ 2 =110 мВ потенциал действия
φ 4=-30 мВ запаздывающий потенциал.
Представим схему развития и распространения потенциала действия по аксону нервной клетки:
1. Состояние покоя.

Положительные и отрицательные заряды равномерно распределены вдоль аксона. Потенциал покояφ 1 = -70 мВ.
2. Приложен стимул.
 Происходит увеличение проницаемости мембраны для ионов натрия, примерно в 5000 раз. Поток ионов натрия из межклеточного пространства перекрывает отрицательный потенциал клетки и разрушает локальный положительный потенциал. Этот потенциал возникает в доли секунды и вырастает до φ 2 =40мВ и называется потенциалом действияφ 3 =110 мВ
Происходит увеличение проницаемости мембраны для ионов натрия, примерно в 5000 раз. Поток ионов натрия из межклеточного пространства перекрывает отрицательный потенциал клетки и разрушает локальный положительный потенциал. Этот потенциал возникает в доли секунды и вырастает до φ 2 =40мВ и называется потенциалом действияφ 3 =110 мВ
3. Распространение потенциала действия.
 Перемена мембранного потенциала называется деполяризацией. Эффект деполяризации, возникший в одной точке, индуцирует деполяризацию с обеих сторон от возбужденной точки.
Перемена мембранного потенциала называется деполяризацией. Эффект деполяризации, возникший в одной точке, индуцирует деполяризацию с обеих сторон от возбужденной точки.
Возникшее однажды значение потенциала действия распространяется по аксону до тех пор, пока не деполяризуется весь участок.
4. Полная деполяризация участка аксона.
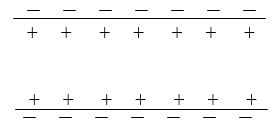 Почти сразу же после деполяризации мембрана приходит в исходное состояние с низкой проницаемости для ионов натрия.
Почти сразу же после деполяризации мембрана приходит в исходное состояние с низкой проницаемости для ионов натрия.
5. Начало реполяризации.
 Происходит резкое увеличение проницаемости мембраны для ионов калия. Ионы калия диффундируют из клетки, и это приводит к понижению потенциала клетки до отрицательных значений.
Происходит резкое увеличение проницаемости мембраны для ионов калия. Ионы калия диффундируют из клетки, и это приводит к понижению потенциала клетки до отрицательных значений.
«Ворота» для калия остаются открытыми в течение всего процесса реполяризации и, следовательно, потенциал падает ниже потенциала покоя и составляет минус 100 мВ. Клетка приходит в исходное состояние за 10 мсек.
Таким образом, формирование потенциала действия обусловлено двумя ионными потоками через мембрану: поток ионов натрия внутрь клетки приводит к перезарядке мембраны, а противоположно направленный поток ионов калия обуславливает восстановление исходного потенциала покоя.
Общее изменение мембранного потенциала при возбуждении складывается из потенциала покоя и мембранного потенциала при возбуждении  . Потенциал действия рассчитывается по уравнению Ходжкина – Хаксли.
. Потенциал действия рассчитывается по уравнению Ходжкина – Хаксли.

В состоянии возбуждения, отвечающим генерации потенциала действия, отношение ионных проницаемостей, следующее:

Потенциал действия функционирует по принципу «все или ничего» т.е. процесс либо происходит до конца, либо не начинается. Если стимул имеет величину, достаточную для запускания потенциала действия, то потенциал имеет одинаковую амплитуду и форму независимо от типа, величины и продолжительности действия стимула.
Скорость, с которой потенциал действия распространяется вдоль нервного волокна, зависит от диаметра волокна и от того покрыт ли аксон слоем миелина (структура из липопротеидов). У небольших волокон без миелина скорость может быть меньше 0,5 м/с, в то время как для больших волокон, покрытых миелином, скорость может быть 130 м/с. Ясно, что нервные импульсы не распространяются подобно электрическим сигналам. Распространение нервного импульса – это сложный электрохимический процесс, детали которого подробно рассматриваются при изучении таких дисциплин, как биохимия, физиология, нервные болезни.

