Синапсы в ЦНС (центральные синапсы) служат и механизмом управления деятельностью нервных клеток.
В каждый момент времени отдельные нейроны, нервные центры и нервная система в целом принимают из окружающей среды от разнообразных рецепторов кожи, фоторецепторов сетчатки, волосковых клеток внутреннего уха, хемо-, механо-, барорецепторов внутренних органов множество сигналов. На основе такой разномодальной информации нейроны (как части нервной регуляторной системы) предпринимают действия, адекватные конкретным обстоятельствам, в которых находится человек и его внутренняя среда. Для этого у отдельных нервных клеток и НЦ имеется ряд механизмов (своеобразных «правил»), с помощью которых анализируются приходящие в нервную систему всевозможные типы сигналов. Постсинаптическая мембрана дендритов, сомы и аксона нервной клетки контактирует с пресинаптическими окончаниями десятков, сотен и тысяч других нейронов.
Поэтому на мембране нервной клетке отдельные входные (сенсорные) химические сигналы постоянно «смешиваются», то есть нейрон их объединяет — интегрирует. Указанное действие нейрона заключается в «принятии» оптимального решения в ответ на множество афферентных сигналов: генерировать на аксонном холмике ПД (нейрон возбуждается) и проводить его далее или игнорировать афферентные входы (нейрон тормозится), в таком случае ПД не возникает.
В процессах интеграции в нервной системе участвуют отдельные нейроны, их объединения — нервные центры, нервные цепи рефлекторные дуги, нейронные сети. При этом не следует думать, что рассматриваемые явления могут проистекать изолированно только на уровне одного нейрона или их объединений и т.д. Иными словами, интеграция осуществляется в нервной системе совокупно на многих ее уровнях. Процесс интеграции в нервной системе является способом передачи и обработки химических и электрических сигналов взаимодействующих друг с другом нервных клеток.
Для осуществления интегративной функции нервные клетки используют возбуждающие и тормозные синапсы.
Синапсы осуществляют интеграцию химических сигналов, поступающих на отдельные нервные клетки.
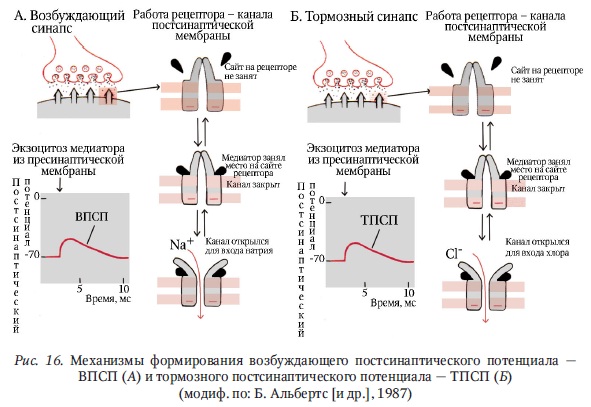
В ЦНС встречаются два вида синапсов между нервными клетками: возбуждающие и тормозные. Потенциалы, возникающие на постсинаптической мембране синапса центральной нервной системы, имеют градуальный характер. Так,возбуждающие постсинаптические потенциалы (ВПСП) в отдельных синапсахЦНС почти никогда не превышают порогового уровня (сравни с НМС). В связис этим для генерации нейроном ПД (распространяющегося возбуждения) необходима одновременная активация нескольких синапсов (сравни с НМС).
В отличие от нервно-мышечных синапсов в синапсах нервной системы работает множество медиаторных систем. В центральных синапсах по сравнению с нервно-мышечными имеются отличия и в структуре постсинаптических рецепторов.
Поясним это на примере: ацетилхолиновый рецептор на постсинаптической мембране мышечного волокна представляет собой натриевый канал. Этот канал открывается под влиянием взаимодействия ацетилхолина с рецепторным сайтом на трансмембранном белке. После чего мембрана миоцита деполяризуется до порогового уровня и возникает ПД. Иными словами, ацетилхолиновый[1] рецептор в нервно-мышечном синапсе является возбуждающим и вызывает возбуждающие постсинаптические потенциалы путем открытия натриевого канала.
В синапсах ЦНС, кроме ацетилхолина, обнаружено большое число других нейромедиаторов. Влияние нейромедиатора определяется спецификой канала на постсинаптической мембране. Специфичность зависит от того, какие ионы пропускает этот канал при взаимодействии рецепторного сайта на мембране нейрона с нейромедиатором. Причем каждый тип нейротрансмиттера управляет определенными ионными каналами. В результате связи нейротрансмиттеров с участком на рецепторах постсинаптических мембран открываются каналы для входа положительных или малых отрицательных ионов, например, Na+, K+, Са2+, Cl- (рис. 16).