




Воздействуя на молочнокислые бактерии различными мутагенными факторами, можно быстрее получить новые штаммы микроорганизмов с требуемыми свойствами [33, 280, 296, 608, 695 и др.].
Повышать частоту мутаций по сравнению со спонтанным фоном, т.е. индуцировать их, могут физические, химические и биологические факторы, действующие на генетический материал клетки. Физические факторы – это, прежде всего, коротковолновое излучение (УФ-излучение и рентгеновские лучи), гамма-лучи.
К химическим мутагенам относятся аналоги оснований, производные акридина, алкилирующие и дезаминирующие агенты. Биологические факторы – это в первую очередь мигрирующие элементы (транспозоны и IS-элементы).
Мутации, независимо от того, имеют ли они спонтанное происхождение или индуцированы каким-либо мутагеном, по характеру перестроек, происшедших в ДНК, можно разделить на мутации, состоящие в изменении одного нуклеотидного остатка молекулы ДНК, так называемые точковые мутации, и мутации, при которых наблюдается изменение участка молекулы ДНК размером больше одного нуклеотида. Точковые мутации, в свою очередь, могут быть разделены на несколько классов в зависимости от того, какие конкретно химические перестройки происходят в молекуле ДНК в рамках одного нуклеотидного остатка: замена, вставка или выпадение. К мутациям, затрагивающим сегмент бактериальной хромосомы, ведут выпадение нескольких оснований или даже генов, перемещение их в пределах одной хромосомы, умножение или удвоение части хромосомы.
Частым типом структурных повреждений ДНК, вызываемых УФ-излучением, является образование пиримидиновых димеров в результате ковалентного связывания соседних пиримидиновых оснований. Реже УФ вызывает разрыв водородных связей, образование межцепочечных поперечных сшивок и поперечных сшивок между ДНК и белком. Ионизирующие излучения всех видов вызывают главным образом одноцепочечные разрывы в ДНК; разрывов, поражающих обе цепи, обычно на порядок меньше. Различные химические мутагены индуцируют образование внутрицепочечных и межцепочечных поперечных сшивок и одноцепочечные разрывы ДНК.

| Пиримидиновый димер – дефект ДНК, возникающий в результате образования ковалентной связи между двумя соседними пиримидиновыми основаниями (тимидином или цитозином) под действием ультрафиолетовых лучей. Ультрафиолетовые лучи вызывают разрыв двойной связи и образование в этом месте ковалентной связи между двумя нуклеотидами. Образование димера приводит к нарушению транскрипции ДНК на данном участке и возникновению мутаций. |
| Р и с. 2.1. Образование пиримидинового димера [Пиримидиновый] |
Наследственная информация у бактерий хранится в форме последовательности нуклеотидов ДНК [123] хромосомы и плазмид.
Размеры бактериальной хромосомы у молочнокислых микроорганизмов составляют 108 – 109 Д. Генетическая информация в бактериях может содержаться во внеядерных (внехромосомных) молекулах ДНК, представленных плазмидами, транспозонами и IS-элементами.

| Плазмиды – дополнительные факторы наследственности, расположенные в клетках вне хромосом и представляющие собой кольцевые (замкнутые) или линейные молекулы ДНК. Плазмиды выполняют регуляторные и кодирующие функции. Первые направлены на компенсацию метаболических дефектов, вторые вносят в бактерию информацию о новых признаках. Как составляющая часть генетического материала бактерии плазмиды играют важную роль в ее жизнедеятельности, детерминируя такие характеристики, как способность продуцировать экзотоксины, ферменты или бактериоцины, устойчивость к лекарственным препаратам и т.д. |
| Р и с. 2.2. Факторы наследственности микроорганизма: 1 – хромосома, 2 – плазмиды |
Плазмиды бактерий представляют собой двунитевые молекулы ДНК размером от 106 до 108 Да, несущие от 40 до 50 генов. Они не считаются жизненно необходимыми, т.к. не кодируют информацию о синтезе ферментов, участвующих в основном метаболизме бактериальной клетки. Благодаря современным достижениям генетики известно, что количество плазмид в микроорганизмах может быть от 0 до 200. Штаммы молочнокислых микроорганизмов содержат наборы плазмид, включающие от 0 до 12 (обычно 4–7) кольцевых ковалентнозамкнутых молекул ДНК массой от 1 до 80 МДа.
Транспозоны и IS-элементы также могут содержаться в бактериальных клетках молочнокислых микроорганизмов [ 695, 707 и др. ].
Транспозоны – внехромосомные генетические элементы (транспозиционные элементы, прыгающие гены, мигрирующие элементы) – последовательности ДНК, которые могут перемещаться с места на место в пределах генома клетки. Транспозоны устроены более сложно, чем IS-элементы: в них включены некоторые гены, не имеющие отношения к процессу транспозиции. Известны транспозоны, содержащие гены устойчивости к антибиотикам, сульфамидам, ионам тяжелых металлов и другим ингибиторам. Транспозоны – это линейные молекулы двунитевой ДНК, размеры которых колеблются 1–6 т.п.н. Мигрирующие элементы могут встраиваться в разные участки бактериальной хромосомы или переходить с бактериальной хромосомы на плазмиду; репликация этих элементов осуществляется под контролем тех же механизмов, что и у соответствующей хромосомы или плазмиды. Частота переносов (транспозиций) мигрирующих элементов колеблется от 10–4 до 10–7.
IS-элементы (от англ. «insertion sequences») – инсерционные (вставочные) последовательности – входят, как правило, в состав хромосом, но способны переходить из хромосомы в плазмиду, поэтому относятся к внехромосомным генетическим элементам. IS-элементы – это линейные молекулы двунитевой ДНК, размеры которых, как правило, до 1 т.п.н.
IS-элементы содержат информацию, необходимую только для их переноса внутри клетки, никаких выявляемых признаков в них не закодировано. Для переноса мигрирующих элементов между клетками нужны переносчики, которыми могут быть определенные плазмиды или фаги. Встраивание мигрирующих элементов в бактериальную хромосому оказывает мутагенное действие, так как при этом происходит включение фрагмента ДНК, приводящее к изменению порядка расположения нуклеотидов в триплете и, как следствие этого, нарушению процесса транскрипции. IS элементы являются простейшей разновидностью транспозиционных элементов и не несут никакой генетической информации, за исключением той, которая необходима для транспозиции
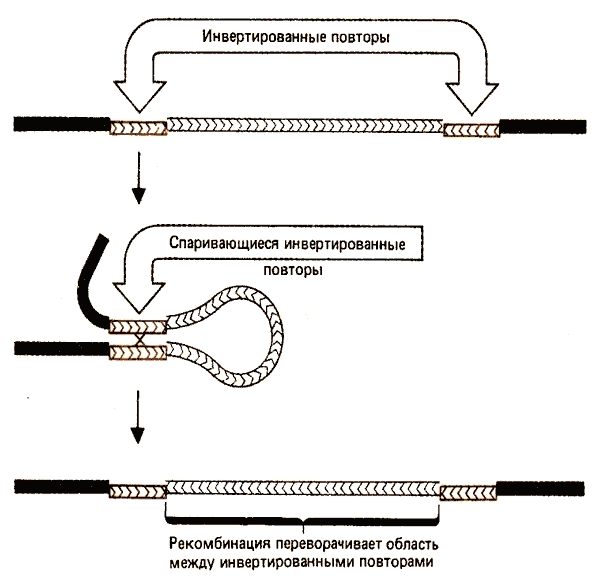
| Р и с. 2.3. Механизм рекомбинации с участием инвертированных повторов IS-элементов [Биология и медицина] IS были обнаружены в бактериях, бактериофагах, плазмидах. Отличительная особенность этих элементов – неспособность к автономной репликации. IS последовательность перемещается на новое место в геноме с использованием фермента транспозазы, ген которой включен в нуклеотидную последовательность IS. |
Еще более широкие перспективы получения производственно-ценных штаммов открывает использование различных способов рекомбинации генов: конъюгации, трансдукции, трансформации. Часто прибегают к трансдукции (перенос гена из одной бактерии в другую посредством бактериофагов), трансформации (перенос ДНК, выделенной из одних клеток, в другие).
Рекомбинация (recombination) – перераспределение генетического материала родителей, приводящее к наследственной комбинативной изменчивости. В общем смысле под рекомбинацией понимают создание новой комбинации генов при соединении генетических факторов родителей. У прокариот рекомбинация осуществляется в процессе конъюгации, трансформации либо трансдукции, у вирусов – при смешанной инфекции. Различают реципрокную (взаимный обмен участками молекулы ДНК), нереципрокную (односторонний перенос участка ДНК); общую (кроссинговер), сайт-спе-цифическую и незаконную рекомбинацию (обмен участками негомологичных хромосом в результате хромосомных перестроек) [411]. Иногда прибегают к трансдукции (перенос гена из одной бактерии в другую посредством бактериофагов), трансформации (перенос ДНК, выделенной из одних клеток, в другие).
При генетических рекомбинациях в хромосому (плазмиду) бактериальной клетки-донора встраивается фрагмент хромосомы или плазмиды клетки-донора. Образующиеся рекомбинанты, в основном, сохраняют генотип микроба-реципиента, приобретаются только отдельные свойства донора-микроба, т.е. происходит нереципроктная рекомбинация.
Конъюга́ция (от лат. Conjugatio – соединение) – однонаправленный перенос части генетического материала (плазмид, бактериальной хромосомы) при непосредственном контакте двух бактериальных клеток. Открыт в 1946 году Дж. Ледербергом и Э. Тайтемом. Имеет большое значение в природе, поскольку способствует обмену полезными признаками при отсутствии истинного полового процесса. Из всех процессов горизонтального переноса генов конъюгация позволяет передавать наибольшее количество генетической информации [224]. В клетке-доноре должна присутствовать конъюгативная (трансмиссивная) плазмида. Первой из них была открыта F-плазмида-эписома (способная встраиваться в бактериальную хромосому), длиной около 100 тыс. пар оснований. Плазмида несет гены, кодирующие ряд функций. Одна из них – образование так называемых пилей, отвечающих за приклепление к клетке-реципиенту. Неконъюгамивная плазмида может передаваться в клекту-реципиент с помощью Тра-генов конъюгативной плазмидой.
Таким образом, для молочнокислых микроорганизмов успешно использовали индуцированный мутагенез, различные способы рекомбинации генов: конъюгацию, трансдукцию, трансформацию и другие генетические процессы. При индуцированном мутагенезе молочнокислых микроорганизмовчаще всего возникает образование пиримидиновых димеров, иногда разрыв водородных связей, одноцепочечные разрывы в ДНК, образование внутрицепочечных и межцепочечных поперечных сшивок ДНК, межцепочечные поперечные сшивоки между ДНК и белком. При конъюгации – нереципроктрая рекомбинация. Частота ингдукции и рекомбмнации обычно составляет 10–9 –10–7. Более высокая эффективность получения мутантов – следствие использования рекомбинации с участием инвертированных повторов IS-элементов, транспозонов.
Важно подчеркнуть, что индуцированный мутагенез и рекомбинация – всего лишь методы более эффективного получения мутаций, или рекомбинаций, соответственно, которые происходят в естественных условиях в природе. Совершенно иным подходом в селекционной работе с микроорганизмами является получение рекомбинантов путем слияния протопластов, которое позволяет объединить генетические материалы и таких микроорганизмов, которые в естественных условиях не скрещиваются, или использование двурепликонных плазмид с геном антибиотикоусточивости.
В этих случаях маркер антибиотикоустойчивости может самопроизвольно конъюгативно передаваться в неродственные микроорганизмы, в т.ч. патогены, что является препятствием для использования такого рекомбинанта в составе пробиотического БП или ПФП.

