




ДЫХАНИЕ РАСТЕНИЙ
Образующиеся в ходе фотосинтеза сахара и другие органические соединения используются клетками растительного организма в качестве питательных веществ. На клеточном уровне этот процесс называется дыханием.
Клеточное дыхание – это окислительный, с участием О2 распад органических питательных веществ, сопровождающийся образованием химически активных метаболитов и освобождением энергии, которые используются клетками для процессов жизнедеятельности.
Суммарное уравнение дыхания:
 С6Н12О6 + О2 6СО2 + 6Н2О + 2875 кДж/моль
С6Н12О6 + О2 6СО2 + 6Н2О + 2875 кДж/моль
Значение дыхания не ограничивается тем, что это процесс поставляющий энергию. Дыхание, подобно фотосинтезу, сложный окислительно-восстановительный процесс, идущий через ряд этапов. На его промежуточных стадиях образуются органические соединения (органические кислоты и пентозы), которые затем используются в различных метаболических реакциях. Кроме того, вода, образующаяся при дыхании, в крайних условиях обезвоживания может быть использована растением и предохранить его от гибели. Таким образом, дыхание – центральный метаболический процесс, переплетающийся многочисленными связями с другими процессами обмена.
Ферменты дыхания
С химической точки зрения дыхание – это медленное окисление, связанное с отнятием от субстрата электронов и протонов, а кислород играет роль их конечного акцептора, что отличает процесс дыхания от брожения (анаэробный процесс).
Окисление дыхательного субстрата осуществляется с участием ферментов оксидоредуктаз (I класс)

 ДН2 Е АН2 Д – донор электронов и протонов
ДН2 Е АН2 Д – донор электронов и протонов
А - акцептор
Д ЕН2 А Е - энзим
 Существует 3 группы оксидоредуктаз:
Существует 3 группы оксидоредуктаз:
1. Анаэробные дегидрогеназы (передают электроны различным акцепторам, но не кислороду). В качестве кофермента – NAD+, NADP+. Например: лактатдегидрогеназа, малатдегидрогеназа.
2. Аэробные дегидрогеназы (передают энергию различным акцепторам, в том числе и кислороду). В качестве простетической группы содержат рибофлавин (витамин В2). Различают два кофермента этой группы: FMN или жёлтый дыхательный фермент Варбурга и FAD.
Пример: сукцинатдегидрогеназа. Доноры электронов для аэробных дегидрогеназ – анаэробные дегидрогеназы, а акцепторы – хиноны, цитохромы, кислород.
- Оксидазы (передают электроны только кислороду). При этом образуются вода, перекись водорода и супероксидный анион кислорода – О2-.
 4е Н2О (цитохромоксидаза, полифенолоксидаза)
4е Н2О (цитохромоксидаза, полифенолоксидаза)
АН2
 + 2е Н2О2 (оксидазы аминокислот)
+ 2е Н2О2 (оксидазы аминокислот)
 О2 1е
О2 1е
О2- + Н+ (ферменты типа ксантиноксидазы)
Н2О2 и О2 весьма токсичные, поэтому в клетках быстро трансформируются в воду и кислород.
Среди оксидаз очень важную роль играют железосодержащие ферменты и переносчики, относящиеся к цитохромной системе. В неё входят цитохромы и цитохромоксидаза. Именно они передают электроны от флавопротеинов на кислород.
Все компоненты цитохромной системы содержат железопорфириновую простетическую группу; при переносе электронов железо обратимо восстанавливается и окисляется, приобретая или отдавая электроны и изменяя таким образом свою валентность.
Другие растительные оксидазы (полифенолоксидаза, аскорбатоксидаза) являются медьсодержащими и немитохондриальными (окисляют соответственно фенолы и аскорбиновую кислоту).
К оксидазам относятся также пероксидазы (используют в качестве окислителя перекись водорода) и каталаза (расщепляет перекись водорода на воду и кислород). Простетической группой каталазы и пероксидазы служит гем.
Субстраты дыхания
В первую очередь растения используют в качестве дыхательного материала углеводы. При этом дыхательный коэффициент (молярное отношение СО2, выделившегося в процессе дыхания, к поглощённому за этот же промежуток времени О2) равен единице (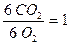 ).
).
При недостатке углеводов могут быть использованы и другие субстраты. Особенно ярко это проявляется на проростках, развивающихся из семян, в которых в качестве запасного питательного вещества содержаться белки или жиры. Так как эти соединения по сравнению с сахарами являются более восстановленными, для их окисления требуется больше кислорода и дыхательный коэффициент становится меньше единицы. При этом жиры предварительно расщепляются до глицерина и жирных кислот, которые могут быть превращены в углеводы через глиоксилатный цикл. Использованию же белков в качестве субстрата дыхания предшествует их расщепление до аминокислот.
ПУТИ ДЫХАТЕЛЬНОГО ОБМЕНА
Существует два основных пути превращения дыхательного субстрата или окисления углеводов:
1. Дихотомический: гликолиз + цикл Кребса;
2. Апотомический: пентозофосфатный путь
Относительная роль этих путей дыхания может меняться в зависимости от типа растений, возраста, фазы развития, а также в зависимости от условий внешней среды (например, дыхание осуществляется в диапазоне температур -50….+50 0С).
Рассмотрим оба пути:
Гликолиз – процесс анаэробного распада глюкозы, идущий с освобождением энергии, конечным продуктом которого является пировиноградная кислота (ПВК).
Реакции гликолиза протекают в растворимой части цитоплазмы (цитозоле), где гликолитические ферменты организованы в мультиэнзимные комплексы с участием актиновых филаментов цитоскелета, и в хлоропластах.
Цепь реакций, составляющих путь гликолиза можно разбить на 3 этапа:
1. 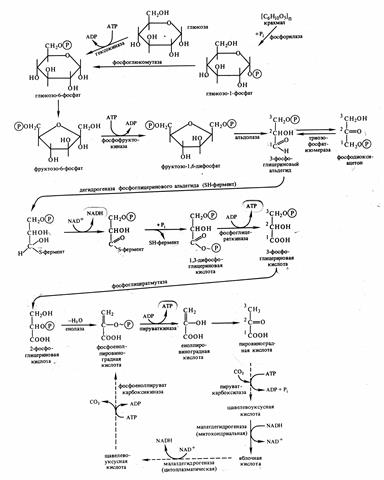
Подготовительный – фосфорилирование гексозы и её расщепление на 2 фосфотриозы (затрачивается 2 АТФ).
2. Первое субстратное фосфорилирование (синтез 2 АТФ за счёт окисления 2 молекул фосфоглицеринового альдегида (ФГА) до 2 молекул фосфоглицериновой кислоты (ФГК), при этом также образуется 2 NADH 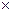 H+).
H+).
3. Второе субстратное фосфорилирование( синтез 2 АТФ за счёт окисления 2 молекул ФГК до ПВК).
Таким образом, суммарное уравнение гликолиза выглядит следующим образом:
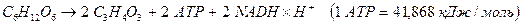
Функции гликолиза в клетке:
1. Связь между дыхательными субстратами и циклом Кребса.
2. Источник АТР и NADH H+.
3. Некоторые его интермедиаты используются для синтетических процессов в клетке (например, ФЕП необходима для образования фенольных соединений и лигнина).
4. В хлоропластах запасённый крахмал метаболизируется в триозы, которые затем транспортируются из него.
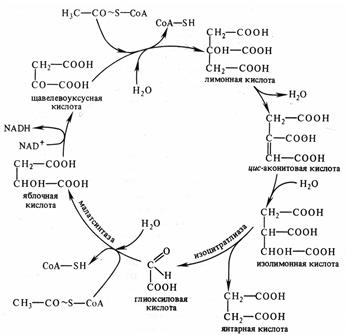
Цикл Кребса. В анаэробных условиях ПВК подвергается различным типам брожения. В присутствии же достаточного количества кислорода пируват полностью окисляется до углекислого газа и воды в цикле Кребса. Все участники этого процесса локализованы в матриксе или во внутренней мембране митохондрий.
Первым этапом окислительного расщепления ПВК является процесс образования активного ацетила в ходе окислительного декарбоксилирования при участии пируватдегидрогеназного мультиферментного комплекса (3 фермента и 5 коферментов).
В результате образуется ацетил коэнзим А:
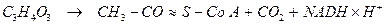
(ацетил – Со А)
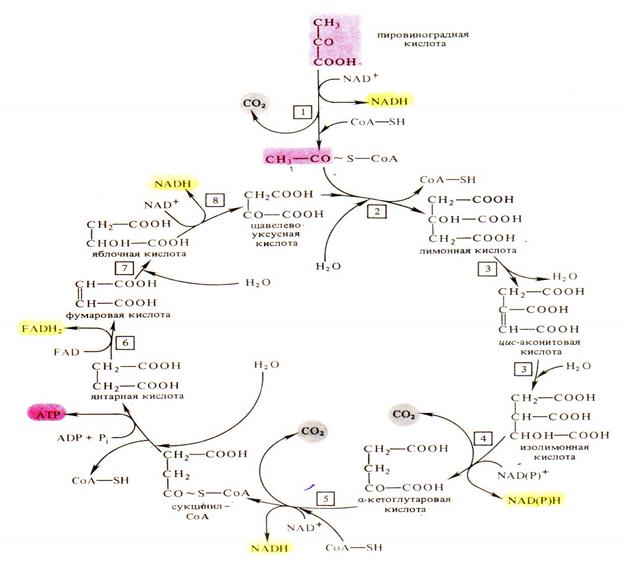
Дальнейшее окисление ацетил - Со А осуществляется в ходе циклического процесса (цикла Кребса) начиная со взаимодействия со щавелевоуксусной кислотой (ЩУК):
В результате одного оборота цикла:
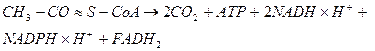
Сумарне рівняння аеробного окислення:
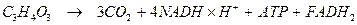
Сумарне рівняння гліколізу і циклу Кребса:
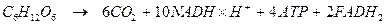
Глиоксилатный цикл (в животных клетках отсутствует). Можно рассматривать как модификацию цикла Кребса. Он активно функционирует в прорастающих семенах масличных растений, где запасные жиры превращаются в сахара (глюконеогенез). Локализован не в митохондриях, а в специализированных микротельцах – глиоксисомах.
В отличие от цикла Кребса, в каждом обороте участвует не одна, а две молекулы ацетил-СоА А (образуется при β-окислении жирных кислот)и используется для синтеза янтарной кислоты, которая выходит из глиоксисом, превращается в ЩУК и участвует в глюконеогенезе (обращённом гликолизе).
 |
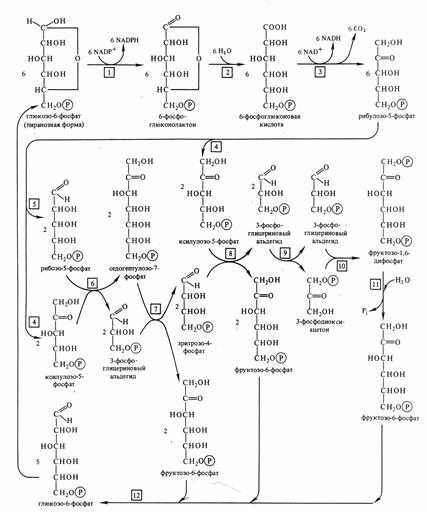
Пентозофосфатный путь окисления (ПФП, пентозный шунт). Окисление глюкозы по этому пути связано с отщеплением от глюкозы первого атома углерода в виде СО2.
Протекает этот процесс в растворимой части цитоплазмы клеток и в хлоропластах. ПФП дыхания особенно активен в тех клетках и тканях растений, в которых интенсивно идут синтетические процессы, такие, как синтез липидных компонентов мембран, нуклеиновых кислот, клеточных стенок, фенольных соединений.
В ПФП выделяют 2 этапа:
1) окисление глюкозы до рибулозо-5-фосфата с образованием 2 NADPH*H+;
2) рекомбинация сахаров для регенерации исходного субстрата при участии ферментов транскетолаз, трансальдолаз и изомераз.
6С5 = 5С6
Суммарное уравнение ПФП выглядит таким образом:
6С6Н12О6 = 5С6Н12О6 + 6СО2 + 12 NADPH*H+
Основное значение ПФП не энергетическое, а пластическое:
1 - NADPH*H+, образующийся не в митохондриях, а в цитоплазме, используются главным образом в различных синтетических процессах (так как в отличие от NAD+, который присутствует в клетках в основном в более устойчивой окисленной форме, NADPH*H+ - в восстановленной).
2 – В ходе ПФП синтезируются пентозы, входящие в состав нуклеиновых кислот и нуклеотидов (ATP,GTP,UTP, NAD,FAD,коэнзим А тоже являются нуклеотидами, в состав которых входит рибоза).
3 – Углеводы С3 – С7 необходимы для синтеза различных веществ. Например, из эритрозы синтезируется шикимовая кислота – предшественник многих ароматических соединений, таких как аминокислоты, витамины, дубильные и ростовые вещества, лигнин клеточных стенок и др.
4 – Компоненты ПФП принимают участие в темновой фиксации СО2. Только 2 из 15 реакций цикла Кальвина специфичны для фотосинтеза, остальные участвуют в ПФП.
5 – Триозы С3 из ПФП могут превращаться в 3-ФГК и участвовать в гликолизе.
 |
Активность ПФП в норме в общем дыхательном обмене составляет 10-40%. Активность увеличивается при неблагоприятных условиях: засухе, калийном голодании, инфекции, затенении, засолении, старении.
Дыхательная электронтранспортная цепь и окислительное фосфорилирование.
Цикл Кребса, глиоксилатный и ПФП функционируют только в условиях достаточного количества О2. В то же время О2 непосредственно не участвует в реакциях этих циклов. Он необходим для заключительного этапа дыхательного процесса, связанного с окислением накопленных восстановленных коферментов NADH*H+ и FADH2 в дыхательной электронтранспортной цепи (ЭТЦ) митохондрий. С переносом электронов по ЭТЦ сопряжен и синтез АТФ.
Дыхательная ЭТЦ, локализованная во внутренней мембране митохондрий состоит из четырех мультиэнзимных комплексов, в состав которых входят (по мере увеличения окислительно-восстановительного потенциала):
FMN – железосерные белки FeS – убихинон Q – цитохромы (b556,b560,1c,c,a,a3) - О2. При этом электроны от NADH*H+ поступают на FMN, а от FADH2 – непосредственно на убихинон.
Из матрикса митохондрий при транспорте каждой пары электронов от NADH*H+ к О2 в трех участках ЭТЦ через мембрану наружу (в межмембранное пространство) переносятся по крайней мере шесть протонов Н+, при окислении FADH2 – таких участков лишь два.
В результате на мембране создается электрохимический потенциал ионов Н+, включающий химический или осмотический градиент (ΔрН) и электрический градиент. Согласно хемиоосмотичской теории Митчелла такой электрохимический трансмембранный потенциал ионов Н+ и является источником энергии для синтеза АТФ за счет транспорта протонов через протонный канал мембранной АТФ-азы.
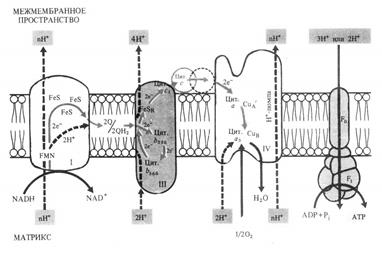 |
АТФ-азный комплекс состоит из фактора сопряжения F1 (белок из 9 субъединиц) и фактора Fо, который перешнуровывет мембрану и служит каналом для транспорта ионов Н+.
Процесс фосфорилирования АДФ с образованием АТФ, сопряженный с переносом электронов при ЭТЦ митохондрий, называется окислительным фосфорилированием.
При окислении NADH*H+ коэффициент фосфорилирования =3, FADH2= 2, то есть энергии хватает на синтез соответственно 3 и 2 молекул АТФ.
Таким образом можно рассчитать общий энергетический выход (в молекулах АЬФ) окисления глюкозы:
С6Н12О6 = 6СО2 + 4 АТФ + 10 NADH*H+ + 2 FADH2.
При этом 10 NADH*H+ = 30 АТФ, а 2 FADH2 = 4 АТФ. В итоге имеем 4 +30+4 =38 АТФ = 380 ккал/моль = 1591 кДж/моль энергии.

