




Нуклеопротеїни (або нуклеопротеїди) — складні білки, комплекси нуклеїнових кислот (ДНК або РНК) з білками. Залежно від типу нуклеїнової кислоти поділяються на дезоксирибонуклеопротеїн та рибонуклеопротеїни. До нуклеопротеїнів відносяться стійкі комплекси нуклеїнових кислот з білками, що тривалий час існують в клітці у складі органел або структурних елементів клітини на відміну від різноманітних короткоживучих проміжних комплексів білок —нуклеїнова кислота (комплекси нуклеїнових кислот зферментами —синтетазами і гідролазами — при синтезі і деградації нуклеїнових кислот, комплекси нуклеїнових кислот з регуляторними білками тощо).
Залежно від типу складових нуклеопротеїдних комплексів нуклеїнових кислот розрізняють рибонуклеопротеїни і дезоксирибонуклеопротеїни.
Стійкість нуклеопротеїдних комплексів забезпечується нековалентною взаємодією. У різних нуклеопротеїдів для забезпечення стабільності комплексу вносять внесок різні типи взаємодій, при цьому нуклеїново-білкові взаємодії можуть бути специфічними і неспецифічними. У разі специфічної взаємодії певна ділянка білка пов'язана із специфічною (комплементарною) нуклеотидною послідовністю, в цьому випадку внесок водневих зв'язків, що утворюються між нуклеотидними іамінокислотними залишками завдяки просторовій взаємній відповідності фрагментів, максимальний. У разі неспецифічної взаємодії основний внесок у стабільність комплексу вносить електростатична взаємодія негативно заряджених фосфатних груп поліаніона нуклеїнової кислоти з позитивно зарядженими амінокислотними залишками білка.
Прикладом специфічної взаємодії можуть служити нуклеопротеїдні комплексирРНК — субодиниці рибосом; неспецифічна електростатична взаємодія характерна для хромосомних комплексів ДНК — хроматіну з гістонами і комплексів ДНК-протаміни головок сперматозоїдів деяких тварин.
Нуклеопротеїни дисоціюють на білки і нуклеїнові кислоти при дії агентів, що руйнують або ослабляють нековалентні зв'язки:
· підвищені концентрації солей або сечовини, що збільшують іонну силу розчину
· іоногені поверхнево-активні речовини
· деякі полярні органічні сполуки (формамід і діметилформамід, фенол тощо).
Деякі нуклеопротеїни (рибосомні субчастинки, нуклеокапсиди вірусів) мають здатність самозбирання, тобто формування, за відповідних умов, нуклеопротеїдів in vitro без участі клітинних структур або агентів; таке самозбирання можливе у разі специфічних нуклеїново-білкових взаємодій (нуклеїново-білковому розпізнаванні). У будь-якому випадку, при утворенні нуклеопротеїдів відбуваються істотні конформаційні зміни нуклеїнових кислот і, в деяких випадках, білків, що утворюють нуклеопротеїдний комплекс.
Дезоксирибонуклеопротеїди
· Хроматин — комплекс ДНК з гістонами в клітинах еукаріотів. За рахунок електростатичної взаємодії нитка ДНК здійснює подвійний оберт навколо октамеру гістонного комплексу H2a, H2b, H3 і H4, утворюючи нуклеосоми, сполучені ниткою ДНК. При приєднанні до комплексу гістону H1 шість нуклеосом утворюють кільцеподібний комплекс, в результаті відбувається конденсація хроматину з утворенням фібрілярної структури, яка далі при приєднанні топоізомерази II і ряду допоміжних білків здатна конденсуватися в гетерохроматин. ДНК, зв'язана в такому нуклеопротеїдному комплексі, не транскрибуєтся.
· Окремим важливим класом дезоксирибонуклеопротеїдів є вірусні нуклеопротеїни. Для реплікації генетичного матеріалу ДНК-вмісних вірусів необхідне перенесення вірусної ДНК в ядро клітини, такий транспорт і проникнення в ядро здійснюються у вигляді нуклеопротеїдних комплексів, білки яких несуть специфічні ділянки — сигнали ядерної локалізації (Nuclear Localization Signal, NLS), що забезпечують транспорт через ядерні пори.
Рибонуклеопротеїни
У клітинах в найбільших кількостях містяться два класи рибонуклеопротеїдів:
· Нуклеопротеїдні комплекси рибосомних РНП (рРНП) — субодиниці рибосом — органел, на яких відбувається трансляція мРНК. Рибосоми є агрегатами з двох різних рРНП-субодиниць.
· Малі ядерні рибонуклеопротеїни (мяРНП) — нуклеопротеїдні комплекси малих ядерних РНК, що є субодиницямисплайсосом (учасників сплайсингу ядерних транскриптів — попередників зрілих мРНК).
· Нуклеопротеїдні комплекси мРНК — матричні рибонуклеопротеїни (мРНП), також відомі як інформосоми. Біологічна роль мРНП вельми різноманітна: вони ймовірно беруть участь у транспорті мРНК, стабілізації (захисту від деградації при транспорті) і регуляції трансляції. мРНП також є хімічно найрізноманітнішим класом нуклеопротеїдів і їхня різноманітність визначається транскриптомом, тобто сукупністю мРНК, що синтезуються в клітині.
Первинна структура нуклеїнових кислот
Нуклеїнові кислоти – це біополімери, які складаються з нуклеотидів, з’єднаних фосфодіефірним зв’язком. Кожний нуклеотид складається із залишків гетероциклічної основи, вуглеводу і фосфорної кислоти.
Одним з найважливіших компонентів нуклеїнових кислот є гетероциклічні основи. Всі вони є похідними піримідину або пурину.
У переважній більшості випадків нуклеїнові кислоти в якості гетероциклічних основ містять урацил (тільки в РНК), тимін (тільки в ДНК) і цитозин, які є похідними піримідину, а також аденін і гуанін, що відносяться до похідних пурину.
Нуклеозиди
У складі нуклеїнових кислот гетероциклічні основи зв’язані з D-рибозою в РНК або з 2-дезокси-D-рибозою в ДНК, утворюючи сполуки, названі відповідно рибонуклеозидами або дезоксирибонуклеозидами. Нуклеозиди – це β-N-пентафуранозиди гетероциклічних основ.
Піримідинові основи з’єднуються з вуглеводом своїми N-1, а пуринові – N-9 атомами азоту. Нуклеозиди, що містять аденін і гуанін, називаються аденозином або дезоксиаденозином і гуанозином або дезоксигуанозином. Нуклеозиди – похідні урацилу і цитозину називаються відповідно уридин або дезоксиуридин та цитидин або дезоксицитидин. Дезоксирибопохідну тиміну прийнято називати тимідином, а рібопохідну – риботииідином.
Нуклеотиди
Третій компонент нуклеїнових кислот – ортофосфорна кислота – утворює складноефірний зв’язок зі спиртовими групами рибози або дезоксирибози. Шляхом розщеплення нуклеїнових кислот в контрольованих умовах вдається виділити складні ефіри нуклеозидів і фосфорної кислоти – нуклеотиди. Назви нуклеотидів формуються від назви гетероциклічної основи, що входить до їх складу, з додаванням слова «кислота»: цитидилова кислота, аденілова кислота і т. д.
Важливу роль в життєдіяльності клітини відіграють складні ефіри поліфосфорних кислот і нуклеозидів. Найбільш широко розповсюджені ефіри, утворені ди- або трифосфатними групами і 5’-ОН-групами нуклеозидів. Вони називаються нуклеозидді або нуклеозидтрифосфатами.
Рідкісні (мінорні) компоненти нуклеїнових кислот
Крім основних компонентів, до складу нуклеїнових кислот можуть входити нуклеозиди з незвичайними гетероциклічними основами або з модифікованим вуглеводним залишком.
Рідкісними компонентами ДНК є метиловані основи: 5-метилцитозин, б-N-метиладенін і деякі інші. У ДНК деяких бактеріофагів замість цитозину є 5-гідроксиметилцитозин і його глюкозовані похідні.
РНК також містять рідкісні компоненти. Вони більше зустрічаються в тРНК, менше – в рибосомних і ядерних РНК. Найбільш поширені похідні звичайних основ (цитозину, урацилу, аденіну та гуаніну) із замісниками в гетероциклічному ядрі або в екзоциклічних аміногрупах. Відомі деякі похідні урацилу.
Дуже часто в тРНК зустрічається продукт дезамінування аденозину – інозин.
Оліго- і полінуклеотиди
Олігонуклеотидами називають полімери, в яких кілька нуклеозидів (до 20) з’єднані один з одним фосфодиефірними зв’язками. Довші ланцюги називають полінуклеотидами.
Різні оліго- і полінуклеотиди відрізняються один від одного за вмістом нуклеотидів кожного типу. Виражені у відсотках відносні кількості мономерних ланок називають нуклеотидним складом, а їх послідовність – первинною структурою нуклеїнової кислоти.
При визначення хімічної будови нуклеозидів, нуклеотидів і полінуклеотидів визначаються тип гетероциклічної основи, природа вуглеводного компонента та місце приєднання його до основи, конфігурація глікозидного центру. Встановлюють також число та місце приєднання залишків фосфорної кислоти.
Гетероциклічні основи у складі нуклеїнових кислот ідентифікуються після жорсткого кислотного гідролізу порівнянням їх хроматографічних характеристик, електрофоретичної рухливості, а також УФ-, ІЧ- та ЯМР-спектрів.
Ідентифікація вуглеводних компонентів також проводиться після кислотного гідролізу нуклеїнових кислот. При цьому з пуринових або попередньо гідрованих за 5, 6-подвійним зв’язком піримідинових рібонуклеотидів утворюється D-рибоза. Основи дезоксирибонуклеотидів відщеплюються в значно м’якших умовах, ніж рибонуклеотиди, але внаслідок нестабільності утворена дезоксирибоза перетворюється у левулінову кислоту. При надзвичайно м’якій обробці кислотою пуринових або відновлених піримідинових дезоксирибонуклеотидів вдається виділити дезоксирибозу або її похідні, які легко ідентифікувати.
Спочатку серед нуклеїнових кислот не було виявлено розгалужених структур, що дозволяло вважати їх виключно лінійними полімерами. Проте згодом була показана можливість існування і розгалужених структур РНК.
Визначення первинної структури
Загальні принципи розшифровки первинної структури нуклеїнових кислот ті ж, що і у випадку інших біополімерів. Полінуклеотидний ланцюг розщеплюється за допомогою ферментів і хімічних агентів, що володіють підвищеною селектвністю, на фрагменти, які дешифруються специфічними методами, і потім реконструюється весь ланцюг.
Знання первинної структури нуклеїнових кислот необхідне для з’ясування зв’язку між їх будовою і біологічною функцією і в кінцевому підсумку – для розуміння механізму їх біологічної дії.
Просторова структура нуклеїнових кислот
Якщо мономерною одиницею нуклеїнових кислот вважати нуклеотидну ланку, то для опису її конформації слід враховувати можливість обертання навколо шести зв’язків. Крім того, слід брати до уваги положення основи щодо вуглеводної частини, яке визначається обертанням навколо N-глікозидного зв’язку, і конформаційні зміни фуранозного циклу. Все це робить конформаційний аналіз нуклеїнових кислот вельми складним.
Конформації нуклеїнових кислот. Двоспіральні полінуклеотиди
У більшості випадків ДНК існує у вигляді «подвійної спіралі».Її основні характеристики зводяться до наступного.Дві полідезоксирибонуклеотидні ланцюги з’єднані один з одним за допомогою водневих зв’язків і утворюють правогвинтову спіраль навколо загальної осі.Ланцюги подвійної спіралі антипаралельні і комплементарні, тобто утворення водневих зв’язків (поперечних) завжди відбувається між основами C і G або А і Т.
Гетероциклічні основи повернені всередину подвійної спіралі, і їх площини приблизно перпендикулярні до її осі. Подвійна спіраль в першому наближенні регулярна, тобто всі її витки мають практично однакові розміри, на кожний виток припадає однакове число пар основ і кожна пара основ повернута відносно іншої на один і той же кут. Із зовнішнього боку спіралі знаходиться гідрофільний вуглеводно-фосфатний залишок.
Комплементарність двох ланцюгів приводить до дуже простого принципу подвоєння генів, або реплікації. Для цього достатньо, щоб ланцюги ДНК розділилися і на кожному з них був синтезований новий комплементарний ланцюг. В результаті утворюються дві дочірні молекули ДНК, ідентичні до вихідної.
Якісного уявлення про структуру ДНК недостатньо для розуміння багатьох спектрів її функціонування, зокрема механізмів взаємодії з білками. Для цього необхідна інформація про деталі структури і можливості її зміни під дією різних факторів.
Циклічні ДНК і суперспіралізація
Багато ДНК з двома ланцюгами в природі є циклічними: плазміди, ДНК мітохондрій і хлоропластів, ДНК багатьох вірусів і бактерій. Такі ДНК, як правило, існують у суперспіральному стані. При цьому подвійна спіраль закручується сама на себе, кількість витків утвореної суперспіралі залежить від зовнішніх умов. Суперспіралізація циклічних ДНК приводить до сильної зміни фізичних властивостей молекули, особливо гідродинамічних і електрофоретичних. У клітинах суперспіралізація здійснюється особливими ферментами, які для бактерій порівняно добре вивчені і називаються ДНК-гіразами (або топоізомеразами II). Інші ферменти – топоізомерази I – можуть зменшувати число супервитків в кільцевих молекулах, даючи набір «ізомерів» з різним числом витків.
Конформації одноланцюгових нуклеїнових кислот
Багато нуклеїнових кислот – більшість РНК і ряд ДНК – існують в одноланцюговій формі. Найбільш добре вивченими одноланцюговими полінуклеотидами є тРНК.
Конформація тРНК
Транспортні РНК виконують в клітинах різноманітні функції. Однак основне їх завдання полягає в здійсненні трансляції. В 1965 році Р. Холлі встановив структуру тРНК з дріжджів. Тоді ж, виходячи з уявлення, що найбільш стабільний стан тРНК відповідає утворенню максимально можливої кількості воднево з’язаних пар основ, а також ґрунтуючись на експериментальних даних щодо нерівномірного гідролізу молекули рибонуклеазами, Р. Холлі запропонував модель вторинної структури тРНК незвичайної форми – з одно- та двохланцюговими ділянками, що чергуються, і назвав цю структуру «лист конюшини».
Структура типу «лист конюшини» пояснювала характерну реакційну здатність нуклеотидних ланок в різних ділянках тРНК у відношенні до хімічних реагентів і до дії рибонуклеаз. Однак гідродинамічні характеристики молекули свідчили про її більш компактну упаковку, яка могла б здійснюватися за рахунок третинної структури. Спосіб утворення третинної структури став зрозумілим після рентгеноструктурного аналізу першої тРНК, яку вдалося отримати в кристалічному. Рентгеноструктурні дослідження були виконані одночасно двома групами. Зрештою були встановлені третинні структури ще декількох тРНК. Третинна структура молекули тРНК нагадує латинську букву L.
Нуклеїнові кислоти - ДНК і РНК - влаштовані за одним принципом. Це складні полімерні молекули, які складаються з безлічі ланок, кожне з яких включає в себе цукор, фосфорну кислоту і азотну основу.
Однак відмінності між ними існують. У цукрі РНК (рибоза) присутній ОН-група, на відміну від цукру ДНК (дезоксирибози). Крім того, ДНК і РНК відрізняються по одному з чотирьох азотистих основ: тимін (Т) в ДНК відрізняється від урацилу (U) в РНК наявністю метильної групи. Молекули РНК виконують ключові функції в ході біосинтезу білка. При «включенні» гена відбувається локальне розплітання спіралі ДНК. Потім з гена, що кодує білкову молекулу, синтезується його РНК-копія. Після ряду «перетворень» вона стає матричної РНК, тобто матрицею для синтезу білка. мРНК переноситься з ядра клітини в цитоплазму, де зв'язується з рибосомами, на яких і «виробляється» білок. Він синтезується з активованих амінокислот, приєднаних до спеціальним транспортним РНК.
Молекули РНК відіграють ключову роль у декодуванні генетичної інформації і біосинтезі білка.
Чим більше заглиблювалися у вивчення різних біосинтетичних процесів, тим частіше виявляли раніше невідомі функції РНК. Виявилося, що крім процесу транскрипції (синтезу РНК шляхом копіювання ділянки ДНК) у ряді випадків, навпаки, може відбуватися синтез ДНК на РНК-матрицях. Цей процес, названий зворотною транскрипцією, використовують в ході свого розвитку багато вірусів, у тому числі сумнозвісні онкогенні віруси і ВІЛ-1, що викликає СНІД.
Таким чином, з'ясувалося, що потік генетичної інформації не є, як спочатку вважалося, односпрямованим - від ДНК до РНК. Роль ДНК як спочатку головного носія генетичної інформації стала піддаватися сумніву. Тим більше що багато вірусів (грипу, кліщового енцефаліту та інші) взагалі не використовують ДНК в якості генетичного матеріалу, їх геном побудований виключно з РНК.
4. Генетичні порушення метаболізму глікогену (глікогенози, аглікогенози)
Спадкові порушення реакцій розщеплення або синтезу глікогену проявляються у вигляді ензимопатій — хвороб накопичення глікогену — глікогенозів та аглікогенозів.
Глікогенози
Глікогенози — спадкові хвороби, молекулярною основою виникнення яких є уроджена недостатність синтезу певних ферментів глікогенолізу, пов’язана з дефектами в генетичній системі клітин. При глікогенозах у внутрішніх органах та тканинах (здебільшого в печінці, м’язах, клітинах крові) спостерігається накопичення аномально надмірної кількості глікогену, іноді зі зміненою молекулярною структурою, який не може використовуватися у метаболічних процесах.
Біохімічні характеристики глікогенозів
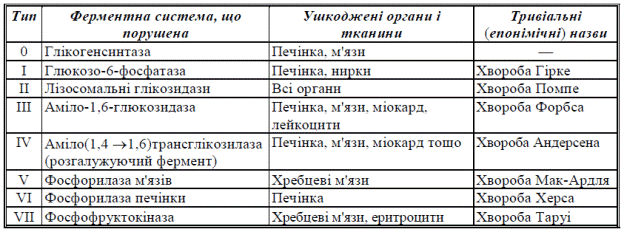
Клінічно глікогенози проявляються важкою гіпоглюкоземією внаслідок нездатності глікогену печінки розщеплюватися з вивільненням молекул глюкози.Глікогенози, при яких ушкоджені ферментні системи мобілізації глікогену печінки, характеризуються збільшенням маси органа, жировою дистрофією гепатоцитів та явищами цирозу. Недостатність ферментних систем глікогенолізу в м’язах супроводжується судомами за умов фізичних навантажень.
Аглікогенози
Аглікогенози — спадкові хвороби накопичення глікогену, молекулярною основою яких є генетичні дефекти, що призводять до порушення утворення ферменту глікогенсинтази. Внаслідок недостатності глікогенсинтази гепатоцити не здатні утворювати резерви глікогену, концентрація якого всередині клітин значно зменшена. Внаслідок відсутності глікогенових резервів хворі при аглікогенозах, як і при глікогенозах, страждають від глибокої гіпоглюкоземії, особливо натщесерце, після значної перерви з часу надходження харчової глюкози. Гіпоглюкоземія при аглікогенозах може супроводжуватися важкою комою внаслідок енергетичного голодування головного мозку. Такі хворі звичайно вмирають у ранньому дитячому віці.

