




Дезоксирибонуклеин қышқылы (ДНҚ) - тірі организмдердегігенетикалық ақпараттың ұрпақтан-ұрпаққа берілуін,сақталуын,дамуы мен қызметін қамтамасыз етуіне жауапты нуклеин қышқылының екі түрінің бірі. ДНҚ-ның клеткадағы басты қызметі - ұзақ мерзімге РНҚ мен белокқа қажетті ақпаратты сақтау. ДНҚ-ның ерекшелiгi. Бiр организмнiң барлық клеткаларындағы ДНҚ молекуласының құрамы, құрылымы бiрдей болады да, жасына, ортадағы жағдайына тәуелдi емес. ДНҚ молекуласының нуклеотидтiк құрамы, құрылымы, тiзбегiндегi нуклеотидтердiң реттелiп орналасуы организмнiң ерекше қасиетiн анықтайды. ДНҚ молекуласының полинуклеотид тiзбегiндегi нуклеотидтердiң ретi – ұрпақтан-ұрпаққа берiлетiн генетикалық мәлiмет. Полинуклеотид тiзбегiндегi нуклеотидтердiң реттелiп орналасуы ДНҚ молекуласының бiрiншi реттiк құрылымы деп аталады. ДНҚ молекуласының екiншi реттiк құрылымын 1953 ж. Уотсон мен Крик анықтады. ДНҚ құрылымының анықталуы ХХ ғасырдағы биологияның ең маңызды жаңалығы деп саналады. Уотсон мен Крик теориясы бойынша екi полинуклеотид тiзбегiнен құралған ДНҚ-ның молекуласы кеңiстiкте оң қос қабат спираль болып табылады. Қос қабат спиральдағы екi тiзбектiң жолдамасы – антипараллель, бiр тiзбектегi нуклеотидтер арасындағы байланыс 3'®5'-бағыттағы қалдықтардан түзiледi, екiншi тiзбектегi нуклеотидтер арасындағы байланыс 5'®3' бағыттағы қалдықтардан түзiледi. Екi полинуклеотидтi тiзбек өзара бұранда сияқты жалғасып, азоттық негiз арқылы байланысады. Гидрофобты азоттық негiздер спиральдiң iшiне орналасқан, ал гидрофильдi пентозды-фосфорлы қалдықтар ДНҚ молекуласының сыртқы жағына қарай бағытталған. Спиральдiң бiр айналымына азоттық негiздiң 10 жұбы келедi. Спиральдiң диаметрi 2 нм болады. Қос қабат спиральдегі азоттық негiздердің қабысуы өте ерекше. Бiр тiзбектегi аденинге – екiншi тiзбектегi тимин, ал гуанинге цитозин қарсы тұрады. Бұл – ДНҚ молекуласының құрылымыныњ өте ерекше маңызды қасиетi. Спиральдағы азоттық негiздердiң осылай орналасуы ДНҚ тiзбегiндегi сєйкестiк-үйлесiмдiлiк (комплементарлық) деп аталады. Қос қабат спиральдi азоттық негiздердiң арасындағы сутектiк байланыс және гидрофобты әрекеттесулер бiрiктiрiп ұстап тұрады. Мұнда аденин мен тиминнiң арасында екi сутектiк байланыс түзiледi, ал гуанин мен цитозиннiң арасында үш сутектiк байланыс түзiледi. Қосақтың әрқайсысында азоттық негiздердiң пентозды-фосфорлы керегесi-мен қосатын гликозидтік байланыстарының арасындаѓы қашықтығы бiрдей – 1,085 нм.
Дезоксирибонуклеин қышқылы (ДНҚ) – барлық тірі клеткалардың негізгі генетикалық материалы болып табылатын күрделі биополимер. ДНҚ-ның негізгі құрылымдық бірлігі – үш бөліктен құралған нуклеотид. Бірінші бөлігі – дезоксирибоза (бескөміртекті қант); екіншісі – пуриндік негіздер: аденин (А) мен гуанин (Г) және пиримидиндік негіздер: тимин (Т) мен цитозин (Ц); үшіншісі – фосфор қышқылының қалдығы. Нуклеин қышқылдарында мономерлік қалдықтар (нуклеотидтер) өзара фосфодиэфирлік байланыспен байланысқан. ДНҚ барлық тірі организмдердің болашақ ұрпағының құрылысы, дамуы және жеке белгілері туралы биол. мәліметті сақтап, оларды жаңадан пайда болатын клеткаларға бұлжытпай «жазу» жүйесінің негізі болып табылады. 1940 жылдың аяғында америкалық биохимик Э.Чаргафф (1905 ж.т.) әр түрлі организмдердің ДНҚ молекуласына талдау жасап, оның құрамындағы А мен Т, Г мен Ц негіздерінің молярлық мөлшері тең екенін көрсетті (бұны Чаргафф ережесі деп атайды). 1952 ж. ағылшын биофизигі М.Уилкинс (1916 ж.т.) және т.б. ғалымдар рентгендік талдау арқылы ДНҚ молекуласы құрылымының спираль бойынша оң жақ оралымын (В – ДНҚ), ал 1979 ж. америкалық ғалым А.Рич (1929 ж.т.) молекула құрылымының сол жақ оралымын (Z – ДНҚ) ашты. Азотты негіздер спираль осіне перпендикуляр түрінде орналасады. ДНҚ-ның үш сатылы құрылымының кеңістіктік моделін алғаш рет 1953 ж. америкалық ғалым Д.Уотсон (1928 ж.т.) мен ағылшын биологы Фрэнсис КрикФ.Крик (1916 ж.т.) жасады. Модель бойынша ДНҚ молекуласы қос тізбектен құрылған. Қос тізбек бір-бірімен азотты негіздер арасында пайда болатын сутекті байланыстар арқылы жалғасады. Бұл қос тізбекті негіздерге комплементарлық (ұқсас) принцип тән, яғни аденинге әдетте тимин, ал гуанинге цитозин сәйкес келеді. ДНҚ-ның бір-біріне қарама-қарсы бағытталған екі спиральді полинуклеотидті тізбегі бір осьті айнала оралып жатады. Уотсон мен Крик моделінің көмегімен ДНҚ-ның өздігінен екі еселену (репликация) қасиеті ашылды. Осы жаңалықтары үшін Уотсонға, Крикке және Уилкинске Нобель сыйлығы берілді (1962). Екі еселену кезінде комплементарлы орналасқан азотты негіздердің сутекті байланысы үзіліп, ДНҚ жіпшелері екіге ажырайды да, екі ұқсас спиральді ДНҚ тізбегі пайда болады. ДНҚ-ның екі еселенуінің мұндай процесі жартылай консервативтік деп аталады, себебі жаңа түзілген ДНҚ молекуласында бір тізбек бұрынғы болады да, екінші тізбек жаңадан түзіледі. Осының нәтижесінде организмнің барлық клеткаларындағы генетик. материал өзгеріссіз қалады. Бұл ғыл. жетістіктер тірі организмнің тұқым қуалаушылығы мен өзгергіштігін молек. деңгейде түсіндіруге жол ашты.
23. Белок биосинтезінің негізгі этаптары. Белок биосинтезі 4 кезеңнен тұрады: 1. амин қышқылдарының активтенуі және аминоацил-тРНҚ түзілуі;2. Белок биосинтездеушікемплекс түзілуі;3 полипептидтік тізбек синтезі (трансляция). 4. Белок конформациясының түзілуі.
1.Амин қышқылы активтенуі АТФ энергиясы есебінен магний иондарының қатысуымен жүреді. Сондай-ақ әр амин қышқылын активтендіретін аминоацил-тРНҚ синтетаза ферменті болады. Активтенген амин қ-лы аденозиннің 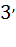 - ОН тобымен тРНҚ – ның акцепторлық шетімен жалғасып, аминоацил-тРНҚ түзеді. Оның 3 учаскесі бар: бірі – тиісті амин қышқ.таниды, екіншісі-тРНҚ-ны, үшіншісі-АТФ-пен жалғасады. 2. иРНҚ рибосоманың кіші суббөлігі 30S – пен жалғасады, бұдан кейін оған алғашқы инициациялаушы аминоацил-тРНҚ қосылады. Осылайша бастапқы комплекс түзіледі. Бастапқы комплекс 50S субббөлікпен қосылып белок синтездеуші комплекс түзеді. 3. Трансляция – белок биосинтезіндегі маңызды процесс. 3-ке бөлінеді.
- ОН тобымен тРНҚ – ның акцепторлық шетімен жалғасып, аминоацил-тРНҚ түзеді. Оның 3 учаскесі бар: бірі – тиісті амин қышқ.таниды, екіншісі-тРНҚ-ны, үшіншісі-АТФ-пен жалғасады. 2. иРНҚ рибосоманың кіші суббөлігі 30S – пен жалғасады, бұдан кейін оған алғашқы инициациялаушы аминоацил-тРНҚ қосылады. Осылайша бастапқы комплекс түзіледі. Бастапқы комплекс 50S субббөлікпен қосылып белок синтездеуші комплекс түзеді. 3. Трансляция – белок биосинтезіндегі маңызды процесс. 3-ке бөлінеді.
Инициация не полипептидтік тізбек синтезінің басталуы. тРНҚ 30 S субббөлікпенIF 3 факторы әсерімен бірігеді. IF 3 фактор 30S және 50S суббөліктерді біріктірмей тұрады. Бұл 3-еуі бастаушы комплексті құрайды. Одан кейін бастаушы комплекс 50 Sсуббөлікпен байланыысады.IF-2 қатысуымен ГТФ гидролизденеді,нәтижесінде ГДФ және фосфор бөлініп шығады. Активті рибосома 70S түзіліп,белок синтезіне дайын болады. Элонгация-полипептидтік тізбектің ұзаруы-циклды процесс.оОл 3 кезеңнен тұрадвы.1.аминоацил-тРНҚ-ның А-учаскесімен байланысуы,2. Пептидтіек байланыстың түзілуі,3..рибасоманың транслокациясы.Синтезделетін белок молекуласында қанша амин қышқылы кодталады,сонша рет осы 3 цикл қайталана береді. Бірінші кезең процесінде А-учаскеге тиісті аминоацил-тРНҚ орналасып,кодон-антикодон бір-бірін танығаннан кейін екінші кезең басталады.А-учаскеде аминоацил-тРНҚ орын алады,ал П-учаскеде N- форилметионил-тРНҚ бар,екеуінің арасында пептидтік байланыс түзіледі.Оны 50S суббөліктің құрамындағы ерекше фернмент пептидилтрансфераза катализдейді. N-формилметионилдің қалдығы тРНҚ-дан А-учаскедегі аминоацил-тРНҚ-ның амин тобына көшіріледі.А-учаскеде дипептилил –тРНҚ түзіледі.П-учаске тРНҚ- аминоацилсыз бос қалады.
Терминация-пептидтік тізбек ұзаруының соңғы сатысы- биосинтездің аяқталуы.Рибосомалар мРНҚ молекуласының бүкіл ұзына бойынан жүріп өтіп,ондағы терминация сигналына жеткенше,генетикалық информация трансляциясы созыла береді.Полипептид синтезінің аяқталу белгісі УАА, УАГ, УГА сияқта мағынасыз кодондардың біреуі болады. Төртінші кезең Белоктың кеңістіктік құрыламының қалыптасуы.Өсіп келе жатқан полипептид тізбегі біртіндеп бүктеледі,өзінің синтезделуі аяқталғаннан кейін 3 өлшемдік өқрылымға ие болады.Полипептид синтезі аяқталғаннан кейін молекула энергентикалық тұрғыдан ең тиімді күйге көшіп,өзінен-өзі тұрақты да сәйкес конформацияға түседі.Белок конформациясының қалыптасуында амин қышқылдарындағы бүйірлік топтар,олардың полярлығы және полярсыз амин қышқылдарының сан қатынасына және тізбектегі олардың орнына байланысты,қалыптасқан белоктың үшінші реттік құрылымы,олардың ылғалды орнтадағы ерекше беріктігін қамтамасыз етеді.Өйткені белок ылғалды ортпада болады.
24.Про- және эукариот клеткаларының нуклеин қышқылдары. Про және эукариот рибосомалары.
Нуклеин қышқылдары (лат. nucleus — ядро) — құрамында фосфоры бар биополемерлер. Молекулалары нуклеотидтерден тұрады, бір нуклеотидтіқ 5'-фосфор арасындағы эфирлік байланысы мен келесі нуклеотидтің углевод қалдығының 3'-гидроксилі арасы эфир байланысымен нуклеин қышқылдары углеводты-фосфатты қаққасын калайды. Нуклеин қышқылдары жоғарғы полимерлі тізбектері ондаған немесе жүздеген нуклеотидтің қалдықтарынан тұрады. Олардың м. с. 105—1010. Нуклеин қышқылдары тірі жасуша ядросының маңызды құрам бөлігі. нуклеин қышқылдарының құрамына азотты негіздер (пиримидинді, пуринді), фосфор қышқылы және моносахаридтер (рибоза мен дезоксирибоза) кіреді.
Нуклеин қышқылдары (НҚ) рибонуклеин қышқылы (РНҚ) және дезоксирибонуклеин қышқылы (ДНҚ) болып екі үлкен түрге бөлінеді.
ДНҚ молекулалық массалары бірнеше мыңнан ондаған миллионға жетеді.ДНҚ мен РНҚ құрамының айырмашылығы — нқ-ын толық гидролиздеу арқылы анықталды. Оларды гидролиздегенде, әр түрлі заттардың қоспасы түзіледі
НҚ-ң 1-құрылымында мононуклеотидтер белгілі тәртіппен орналасады.
2- құрылымы макромолекулалардың кеңістікте қос шиыршық болып орналасуын көрсетеді. Бұл кезде молекулалар арасында және молекула ішінде сутектік байланыс арқылы әрекеттесу болады.
НҚ-ның макромолекуласы екі полинуклеотидті тізбектен құралады. Олар кеңістікте қос оралма түзеді. Оралманы фосфор қышқылының полиэфирі түзеді, пиримидин және пурин туындыларының жазық молекуласы оралманың ішінде болады, өзара сутектік байланыс арқылы байланыскан.
Сутектік байланыс белгілі бір жұп пиримидин және пурин туындыларының арасында түзіледі. Оларды комплементарлы жұптар деп атайды. Ондай жұптар: тимин (Т) — аденин (А) және цитозин (С) — гуанин (G).
3-құрылымы — ДНҚ мен РНҚ-ның кеңістікте шумақталып орналасуы
Рибосома (rіbes — ағыс және some — дене) — ақуыз синтезін жүзеге асыратын жасуша-ішілік органоид. Рибосомалар - екі орташа: үлкен және кіші өлшем бірлігінен тұрады. Нәруыздар биосинтезін (нәруызда аминқышқылдарды қосады) іске асырады.
Рибосома тірі организмдердің барлығынан дерлік табылған. Әр жасушада шамамен 1000-нан 10000-ға дейін Рибосомалар болады. Шамамен диам. 20 нм-дей. Рибосомалар екі класқа бөлінеді: 70S және 80S (S-седиментация коэфф., Сведберг бірлігі). 70S Рибосома ядросы жоқ прокариоттар жасушаларында, ал 80S Р-лар эукариоттардың цитоплазмасында болады. Химиялық құрамы жағынан Р-лар РНІ-нан және ақуыздардан тұратын нуклеопротеид болып саналады. 70S Bold text Рибосомасының 60 — 65%-ы РНҚ және 35 — 40%-ы ақуыздан, ал 80S Рибосомасының 50%-ы РНҚ-дан, 50%-ы ақуыздан құралады. Құрылымы жағынан Рибосомалар үлкен және кіші суббөліктерден тұрады. Рибосомалар цитоплазмада топтасып (5-70-тен)орналасып, полисомалар (полирибосома) түзеді. Рибонуклеопротеидтен құралған (пен РНҚ) рибосомалар жасуша цитоплазмасында базофилді боялады, олардың құрамында ферменттер де кездеседі.
Олар жасуша цитоплазмасындағы Mg иондарының концентрациясына байланысты диссоциацияланып (молек. ыдырауы) және реассоциацияланып (молекулалардың қайта бірігуі) отырады. Рибосомалар жасушада ақуыз синтезіне қатысатын құраушыларды ұстап тұру, оларды бір-бірімен байланыстыру қызметін атқарады. Эндоплазмалық тордың жарғақшасындағы рибосомалардың бірнешеуі бірігіп полирибосома құрайды. Олар жасуша жасуша тіршілігіне қажетті ақуызды синтездейді. Рибосомадағы ферменттер, магний мен кальцийдің тұздары зат айналым өзгерістеріне қатысады. Рибосомада минералдық элементтерден азот,фосфор, темір және мыс кездеседі. Эндоплазмалық торда және оның жасушаларында орналасқан биосинтез бен ақуызды тасымалдайтын бірыңғай жүйе болып саналады.Рибосома митохондриялар құрамында да болады. Бос рибосомалар жасушаның өзіне керекті протеиндерді, ал эндоплазмалық тор қабырғасындағы рибосомалар жасушадан сыртқа шығарылатын протеиндерді түзуге қатысады.
25.Белок синтезіндегі транскрипция кезеңі. Генетикалық код, оның қасиеттері. Генетикалық код - нуклейн қышқылдары молекуласындағы тұқым қуалаушы ақпараттың нуклеотидтер тізбегі түріндегі біртұтас жазылу жүйесі.Генетикалық кодтың негізгі қасиеті-әрбір амин қышқылының үш нуклеотидімен жазылуында. ДНҚ-ның нуклеотидтік кодының болуы арқасында, ген белоктағы амин қышқылдарының жүйелігін бақылайды. Бір геннің кодондары бірін-бірі жаппайды.Көптеген амин қышқылының қалдықтары бірнеше кодондармен кодталады, әрбір жеке кодон тек бір амин қышқылының қалдықтарымен кодталады, әмбебаптылық генетикалық код барлық зерттелген организмдерге бірдей.Тірі жасушаларда генетикалық код, яғни генмен кодталатын белок синтезі, екі матрицалық процестер (транскрипция және трансляция) арқылы жүзеге асады.
Белок синтезінің реакциялары рибосомада жүреді. Белок синтезінің реакциялары жүру үшін күрделі катализдік ақпарат кажет. Өсіп келе жатқан полипептидтік тізбек иРНҚ молекуласына жақындасуы керек, ол иРНҚ-дағы келесі кодондар тРНҚ-ның молекуласымен қосылуы үшін қажет. Бұл дегеніміз, полипептидтің өсіп келе жатқан ұшы, әрбір жаңа амин қышқылы қосылған сайын, иРНҚ-ның тізбегінің бойымен дәл үш нуклеотидке жылжиды деген сөз. Белок синтезінің осы және басқа кезеңдері синтездің белок молекуласынан және РНҚ-дан құралған ірі мультиферменттік комплекс рибосомада жүретініне байланысты.Белоктардың синтезделуі негізінен екі кезеңнен тұрады:1.Ядролық кезең немесе транскрипция. Мұнда ДНҚ қос тізбегінің біреуінің комплементарлы көшірмесі болып табылатын и-РНҚ синтезі жүреді. Осы жолмен синтезделген и-РНҚ әрі қарай синтезделетін белоктың негізі болып табылады.
2. Цитоплазмалық кезең яғни трансляция. Цитоплазмада 4 әріптік генетикалық информацияның триплеттік кодтың көмегімен 20 әріптік амин қышқылдарынан тұратын белоктың тізбегіне айналу процесі жүреді.
Бактерияларда транскрипция және трансляция бірімен-бірі ілесіп жүреді, яғни ДНҚ-на тәуелді РНҚ-полимераза и-РНҚ-ның синтезін жүргізіп жатқан кезде, и-РНҚ-ның бір шетінде белок синтезі де басталып жатады. Бактериялардың екінші бір ерекшелігі и-РНҚ-ның тіршілік ету уақыты бірнеше минут қана, сонан соң олар тез нуклеаза ферментінің әсерімен ыдырап кетеді. Транскрипция(лат. transcrіptіo – қайта көшіріп жазу) – тірі жасушалардағы рибонуклеинқышқылының биосинтез процесі. Олдезоксирибонуклеин қышқылы (ДНҚ) матрицасында жүреді.Транскрипция аденин, гуанин, тимин және цитозинніңқайталанбалы тізбегінен тұратын ДНҚ молекуласындағы генетикалық ақпараттың іске асуының бірінші кезеңі. Транскрипция арнайы ДНҚ және РНҚ полимераза ферменті арқылы жүреді. Транскрипция нәтижесінде РНҚ молекуласының полимерлі тізбегі түзіледі. Бұл тізбек ДНҚ молекуласының көшірілген бөлігіне комплементарлы болады. Транскрипция процесінің өнімі әр түрлі қызметтер атқаратын РНҚ молекуласының төрт типінен тұрады: 1.рибосомадағы ақуыз синтезінде матрицаның рөлін атқаратын ақпараттық немесе матрицалық РНҚ; 2.рибосоманың құрылымдық бөлігін құрайтын рибосомалы РНҚ; 3.ақуыз синтезі кезінде генетикалық ақпараттың РНҚ-дағы нуклеотидтік “тілді” аминқышқылдық “тілге” ауыстыруға қатысатын тасымалдаушы РНҚ; 4.ДНҚ молекуласының репликациясы (генетикалық ақпаратты дәл көшіруді және оның ұрпақтан ұрпаққа берілуін қамтамасыз ететін нуклеин қышқылдары макромолекуласының өздігінен жаңғыру процесі) кезінде бастама қызметін атқаратын РНҚ.
Амин қышқылдарының активтенуі.Белок синтезіндегі тРНҚ-ның және аминоацил т-РНҚ синтетазаның рөлі туралы толық мәлімет беріңіз.
Белок биосинтезі 4 кезеңнен тұрады: 1. амин қышқылдарының активтенуі және аминоацил-тРНҚ түзілуі;2. Белок биосинтездеушікемплекс түзілуі;3 полипептидтік тізбек синтезі (трансляция). 4. Белок конформациясының түзілуі.
1.Амин қышқылы активтенуі АТФ энергиясы есебінен магний иондарының қатысуымен жүреді. Сондай-ақ әр амин қышқылын активтендіретін аминоацил-тРНҚ синтетаза ферменті болады. Активтенген амин қ-лы аденозиннің - ОН тобымен тРНҚ – ның акцепторлық шетімен жалғасып, аминоацил-тРНҚ түзеді. Оның 3 учаскесі бар: бірі – тиісті амин қышқ.таниды, екіншісі-тРНҚ-ны, үшіншісі-АТФ-пен жалғасады. 2. иРНҚ рибосоманың кіші суббөлігі 30S – пен жалғасады, бұдан кейін оған алғашқы инициациялаушы аминоацил-тРНҚ қосылады. Осылайша бастапқы комплекс түзіледі. Бастапқы комплекс 50S субббөлікпен қосылып белок синтездеуші комплекс түзеді. 3. Трансляция – белок биосинтезіндегі маңызды процесс. 3-ке бөлінеді.
Инициация не полипептидтік тізбек синтезінің басталуы. тРНҚ 30 S субббөлікпенIF 3 факторы әсерімен бірігеді. IF 3 фактор 30S және 50S суббөліктерді біріктірмей тұрады. Бұл 3-еуі бастаушы комплексті құрайды. Одан кейін бастаушы комплекс 50 Sсуббөлікпен байланыысады.IF-2 қатысуымен ГТФ гидролизденеді,нәтижесінде ГДФ және фосфор бөлініп шығады. Активті рибосома 70S түзіліп,белок синтезіне дайын болады. Элонгация-полипептидтік тізбектің ұзаруы-циклды процесс.оОл 3 кезеңнен тұрадвы.1.аминоацил-тРНҚ-ның А-учаскесімен байланысуы,2. Пептидтіек байланыстың түзілуі,3..рибасоманың транслокациясы.Синтезделетін белок молекуласында қанша амин қышқылы кодталады,сонша рет осы 3 цикл қайталана береді. Бірінші кезең процесінде А-учаскеге тиісті аминоацил-тРНҚ орналасып,кодон-антикодон бір-бірін танығаннан кейін екінші кезең басталады.А-учаскеде аминоацил-тРНҚ орын алады,ал П-учаскеде N- форилметионил-тРНҚ бар,екеуінің арасында пептидтік байланыс түзіледі.Оны 50S суббөліктің құрамындағы ерекше фернмент пептидилтрансфераза катализдейді. N-формилметионилдің қалдығы тРНҚ-дан А-учаскедегі аминоацил-тРНҚ-ның амин тобына көшіріледі.А-учаскеде дипептилил –тРНҚ түзіледі.П-учаске тРНҚ- аминоацилсыз бос қалады.
Терминация-пептидтік тізбек ұзаруының соңғы сатысы- биосинтездің аяқталуы.Рибосомалар мРНҚ молекуласының бүкіл ұзына бойынан жүріп өтіп,ондағы терминация сигналына жеткенше,генетикалық информация трансляциясы созыла береді.Полипептид синтезінің аяқталу белгісі УАА, УАГ, УГА сияқта мағынасыз кодондардың біреуі болады. Төртінші кезең Белоктың кеңістіктік құрыламының қалыптасуы.Өсіп келе жатқан полипептид тізбегі біртіндеп бүктеледі,өзінің синтезделуі аяқталғаннан кейін 3 өлшемдік өқрылымға ие болады.Полипептид синтезі аяқталғаннан кейін молекула энергентикалық тұрғыдан ең тиімді күйге көшіп,өзінен-өзі тұрақты да сәйкес конформацияға түседі.Белок конформациясының қалыптасуында амин қышқылдарындағы бүйірлік топтар,олардың полярлығы және полярсыз амин қышқылдарының сан қатынасына және тізбектегі олардың орнына байланысты,қалыптасқан белоктың үшінші реттік құрылымы,олардың ылғалды орнтадағы ерекше беріктігін қамтамасыз етеді.Өйткені белок ылғалды ортпада болады.
27).Трансляция стадиялары: инициация, элонгация, терминация. Полипептидтік тізбектің түзілуі. Энергия көзі. Белок синтезінің тежегіштері
Белок биосинтезі үшін клеткада келесі процестер іске асуы қажет:
1.Транскрипция- РНҚ – полимераза ферментінің қатысуымен РНҚ-ң синтезі. Бұл процесте РНҚ-ң 3 түрінің алғы заттары түзіледі: матрицалық пре-РНҚ, рибосомалық пре-РНҚ, транспорттық пре- РНҚ.
2. РНҚ-транскриптің пісіп жетілуі. Түзілген пре-РНҚ-лардан активті мРНҚ, рРНҚ, тРНҚ-р түзіледі.
3. Белок синтезі мРНҚ-ны матрица ретінде пайдалана отырып іске асады. Бүл процесті трансляция деп атайды. Белок молекуласының синтезі трансляция сатысында іске асады. 4
Трансляция. Белок биосинтезі өте күрделі процесс. Трансляция іске асырылу үшін келесі компонеттер қажет:
1. 20 L-аминқышқылдары- белок молекуласының құрылымдық компоненттері
2. мРНҚ- белок синтезін іске асырушы матрица
3. тРНҚ- аминқышқылдарды рибосомаға жеткізеді.
4. Аминоцил- тРНҚ-синтаза – аминқышқылдардың аденилдену (активтену) реакциясын катализдейді.
5. Магний иондары (Mg2+ ) – аминоцил- тРНҚ-синтазаны, трансляция факторларын ж/е рибосоманың конформациясын тұрақтандыру үшін
6. Рибосома, 2 суббөлігі- белоктың биосинтезі іске асатын органелла.
7. АТФ,ГТФ- белок биосинтезіне қажетті химиялық энергияның көзі.
28).Белоктар мен амин қышқ. ыдырауы.Переаминдеу,тотыға дезаминдеу,амин қышқылдарының декарбоксилденуі.Аммиактың түзілуі. Организмде аминқышқ.қор болып иналмайды және бос күйінде организмнен бөлініп шықпайды.Түзілген аминқышқ.мөлшері жоғары болғанда,барлығы белок синтезі үшін пайдаланылмайды,артығы ыдырайды.Адам мен сүтқоректі жануарларда аминқышқ. Ыдырауының реакциялары негізінен бауырда өтеді.Аминқышқ. катаболизмі α аминотобының бөлінуінен,дезаминдеу немесе трансаминдеу процесінен басталады.Тотыға дезаминдену арқылы аминқышқ.молекуласынан амин тобы тотығу жолымен аммиак түрінде бөлініп кетеді.Реакцияны флавин тәуелді оксидаза н/е НАД тәуелдідегидрогеназа катализдейдіАминқ. Метаболизмінде ең маңызды фермент глутаматдегидрогеназа.Бұл L-глутамин қышқ. тотыға дезаминденуін катализдейтін дегидгеназа,кофермент НАД+ н/е НАДФ+ ,ортаның Рһ- ы физиологиялық мағынасына тең болғанда активті.Глутамин қышқ. тотыға дезаминденуі нәтижесінде бөлінген аминтобы бірден аммоний ионына айналады,сондықтан тотыға дезаминдену реакциясы деп аталады.Аминқышқ. шамалы мөлшері катаболизм процесінде алдымен декарбоксилдену реакциясына түседі.Декарбоксилдену реакциясында аминқышқ. СО2 түрінде бөлінеді,нәтижесінде аминдер түзіледі.Реакцияны декарбоксилаза ферменті катализдейді.Жануар ұлпаларында тирозин,триптофан,валин,гистидин,глутамин,цистеин,аргинин,орнитин окситриптофан,оксиглутамин қышқылы,α аминомалон қышқылдарының декарбоксилденуі анықталған.Адам мен жануарлар организмінде аминқышқ/ң декарбоксилдену реакция түрлерінің ең маңыздысы –α-карбоксил тоьының бөлінуі.Реакция өнімдері –аминдер.Декарбоксилдену реакциясы қайтымсыз.Аминқ-ң декарбоксилазасы күрделі ферменттер,коферменті пиридоксальфосфат.Ферменттің ерекшелігі апоферментпен белгіленеді.Аминқ-н түзілетін аминдер биологиялық активті,сондықтан биогенді аминдер деп аталады.Гистидин декарбоксилдену реакц-ң нәтижесінде гистаминға айналады.Гистаминнің биологиялық әсері кең.Гистаминнің көп мөлшері асқазанның шырышты қабатында түзіледі.Ол асқазанға пепсин мен тұз қышқ-ң бөлінуін арттырады.Биогенді аминдер –организмнің физиологиялық прцестеріне әртүрлі әсер ететін күшті фармокологиялық активті заттар.Организмде биогенді аминдер шамалы мөлшерде түзіледі.Көп мөлшерде жиналғанда олардың әсерінен әртүрлі патологиялар пайда болады.Организмде биогенді аминдерді усыздандырудыңм ерекше механизмі бар.Мыс:тотыға дезаминдеу механизмі арқылы аминдер альдегидтерге айналады.Альдегидтер тотығадыда органикалық қышқылдар түзіледі.Аминқ-ң дезаминдену нәтижесінде аммиак түзіледі.Аммиак өте улы зат,организмнен шығарылуы қажет.Қанның құрамында аммиактың концентрациясы 40-50мкмоль/л төиен болу керек.Адамда түзілген аммиактың 90% жуығы несепнәр түрінде шығарылады.Оның синтезі бауырда жүзеге асады.
29).Организмдегі аммиактың залалсыздану жолдары.Орнитин цикліндегі мочевинаның түзілуі. Аминқ-ң дезаминдену нәтижесінде аммиак түзіледі.Аммиак өте улы зат,организмнен шығарылуы қажет.Қанның құрамында аммиактың концентрациясы 40-50мкмоль/л төиен болу керек.Бұл деңгейден жоғары болғанда аммиактың зиянды әсерінен мидың қызметі бұзылады да,ессіз күй пайда болады.Амминдік азот әртүрлі заттар т үрінде организмнен шығарылады. Адамда түзілген аммиактың 90% жуығы несепнәр түрінде шығарылады.Оның синтезі бауырда жүзеге асады.10%жуығы глутамин ж/е аспарагин түрінде мөлшерде түзіледі.Көп мөлшерде жиналғанда олардың әсерінен әртүрлі патологиялар пайда болады.Организмде биогенді аминдерді усыздандырудыңм ерекше механизмі бар.Мыс:тотыға дезаминдеу механизмі арқылы аминдер альдегидтерге айналады.Альдегидтер тотығадыда органикалық қышқылдар түзіледі.Аминқ-ң дезаминдену нәтижесінде аммиак түзіледі.Аммиак өте улы зат,организмнен шығарылуы қажет.Қанның құрамында аммиактың концентрациясы 40-50мкмоль/л төиен болу керек.Адамда түзілген аммиактың 90% жуығы несепнәр түрінде шығарылады.Оның синтезі бауырда жүзеге асады. мөлшерде түзіледі.Көп мөлшерде жиналғанда олардың әсерінен әртүрлі патологиялар пайда болады.Организмде биогенді аминдерді усыздандырудыңм ерекше механизмі бар.Мыс:тотыға дезаминдеу механизмі арқылы аминдер альдегидтерге айналады.Альдегидтер тотығадыда органикалық қышқылдар түзіледі.Аминқ-ң дезаминдену нәтижесінде аммиак түзіледі.Аммиак өте улы зат,организмнен шығарылуы қажет.Қанның құрамында аммиактың концентрациясы 40-50мкмоль/л төиен болу керек.Адамда түзілген аммиактың 90% жуығы несепнәр түрінде шығарылады.Оның синтезі бауырда жүзеге асады. шығарылады.Сүтқоректілерде азоттың көбісі организмнен мочевина түрде шығарылады.Мочевина бұл инертті,суда жақсы еритін,усыз зат.Несепнәр түзілетін жүйе орнитин циклі д/аталады.Несепнәр синтезі 5-сатыдан тұрады:1)цитрулиннің синтезі-екі реакция,2)аргининнің синтезі-екі реакция,3)несепнәрдің түзілуі-бір реакция.1-ші кезең:аммиак ж/е СО2байланысады да,карбомоилфосфат түзіледі.КФС 1 синтаза катализдейді.Бұл реакция іске асу үшін энергия қажет,ол АТФ 2 молекуласы ыдырағанда түзіледі.Одан соң карбомоилфосфат ж/е орнитин конденсацияланады да цитруллин түзіледі.Реакция орнитинкарбомоилтрансферазаның қатысуымен іскеи асады,митохондрияда өтеді.2-ші сатысында бауырда және цитоплазмада іске асады.Цитруллин аспартатпен конденсацияланып,аргининосукцинат түзіледі.Реакцияны аргининосукцинаткиназа катализдейді. Аргининосукцинат әрі қарай аргинин ж/е фумар қышқ-на дейін аргининосукцинатлиаза қатысуымен ыдырайды.3-ші сатыда аргинин ыдырап,несепнәр ж/е орнитин түзіледі,аогиназа ферментінің қатысуымен іске асады.
30) Глюконеогенез, сатылары, энергия көзі. Глюкнеог.-бұл көмірсу емес қосылыстардан глюкозаның синтезі. Ол организм көп энергия шығарып ауыр жұмыс атқарған кезде маңызды қызмет атқарады. Адамдар\ы гликоген түріндегі глюк.қоры 1 күнгі зат алмасуға жетеді. Жануарларда глюкоза түзілетін алғы зат-сүт қышқ, пирожүзімқышқ, глицерин, аминқышқ. Глюкозаның синтезінде алғы зат ретінде қызмет атқаратын аминқ.глюкогенді аминқ.р д.а. оларға: глицин, аланин, серин, цестеин, треонин, аспарагин қышқ.тирозин, фенилаланин, изолейцин, метионин аминқышқ.жатады. глюкоза синтезі үшін алғы зат- сүт қышқ.ол бұлшық етте түзіліп бауырға тасымалданады. Глюконеог.шамалы мөлшердебүйректе ішектің шырышты қабатында, жүрек бұлшықетінде, ми клеткаларында жүзеге асады. Глюконео.з клетканың цитозолында өтеді. Бұл процеске қатысатын ферменттер цитоплазмада орналасады. Өсімдікте глюкоза көп мөлшерде фотосинтез процесі арқылы СО2-ден түзіледі. Глюконеог.ң 3 жаңа реакциялары: 1.пируваттың фосфоенолпируватқа айналуы алдымен пируват карбоксилдену реакциясы арқылы оксалоацетатқа айналады. Бұл реакция АТФ-ң 1 молекуласының қатысуымен жүреді. Оксалоацетат карбоксилсіздену реакциясына түседі, ГТФ қатысуымен фосфоенол пируватқа айналады 1-ші реакцияны пируваткарбоксилаза, 2-сін фосфоенолпируваткарбоксикиназа қатысуымен жүреді. 2.фруктозо-1,6- дифосфоттан 1 көміртек атомының фосфаттық эфирі гидролизі нәтижесінде фруктозо-6-фосфат түзіледі. Реакцияны фруктозо-1,6- дифосфотаза катализдейді. 3. Глюкозо-6-фосфаттан глюкозо-6-фосфотаза қатысуымен гидролиз реакциясы арқылы глюкоза түзіледі.
31.Крахмал мен гликогеннің ыдырауы,тыныс алу процестерінің түрлері. Тыныс алу процессінің жалпы схемасын көрсетіңіз. Жасушаларда биологиялық синтез және ыдырау процестері үздіксіз жүріп отырады. Синтез процесі жалпы анаболизм (гр. "ана" — тірі дене), ал ыдырау процесі катаболизм (гр. "ката" — өлі дене) деп аталады. Ыдырау процесі кезінде күрделі заттардан жай заттар түзіліп, энергия бөлініп шығады. Мысалы, қанттар органикалық қышқыл мен спиртке ыдыраса, органикалық қышкылдар өз кезегінде көмірқышқыл газы мен суға ыдырайды. Зат алмасудың ерекше белгісі сол, мұнда анаболизм және катаболизм процестері бірдей уакытта сыртқы ортамен өзара тура байланыста жүреді. Пластикалық алмасу немесе анаболизм дегеніміз — биологиялық синтез реакцияларының жалпы жиынтығы. Пластикалық алмасуда жасушаға сырттан келіп түсетін заттардан жасушаішілік заттар түзіледі. Пластикалық алмасу реакцияларына мыналар жатады:қанттар мен полисахаридтердің синтезі; крахмал және целлюлозаның түзілуі; глицерин мен май кышқылдарынан майдың, органикалық қышқылдардан аминқышқылының, аминқышкылы мен қанттардан нуклеин кышқылдары азоттық негіздерінің синтезделуі.
Углеводтар адам және жануарлар үшін негізгі қоректік зат. Ол тағам рационының 50-70% шамасындай. Қрьпылу ерекшклігіне қарай углеводтарды екі топқа бөледі: 1/крахмал, гликоген, сахароза, лактоза, 2/целлюлоза, маннандар, пектиндер, пентозандар және баскалары. Екінші топка жататын углеводтарды омыртқалылардың бөрі бірдей қорыта алмайды. Оларды күйіс кайыратын және кейбір жануалар ғана микробтық ферментгің көмегімен қорыта алады.Полисахаридтер мен олигосахаридтердің қарапайым қосылыстарға ыдырауының екі типті реакциясы бар, ол гидролиз және фосфоролиз. Гидролиздің мысалы ретінде крахмалдың ыдырауын, фосфоролизге глиогеннің ыдырауын келтіруге болады. Крахмал гидролизін гидролаза класына жататын α-,β-, γ- амилаза ферментгері катализдейді.
Гликогеннің ыдырау реакциясы фосфоролизден басталады, бұл кезде гликогеннің шеткі глокозасы фосфоролиздік жолмен ажырайды да, оған бейорганикалық фосфат қосылады. Бұл реакцияны гликоген -фосфорилаза ферменті катализдейді және глюкоза -1-фосфат түзіледі.Гликолиз синтезін инсулин гармоны ынталандырады.Гликогеннің синтезделуі АТФ энергиясы есебінен бос глюкозаның фосфорлануы арқылы басталады, бүл реакцияны гексокиназа ферменті катализдейді, бүл фермент барлық клеткаларда бар. Глюкоза, глюкозо-6-фосфатқа айналады. Гликоген синтезінің
қортынды кезеңінде гликоген-синтетаза ферменті УДФ-глюкозадан глюкозаны шығарып полисахаридтердің өсіп бара жаткан тізбегіне қосады.Гликолизде ыдырайтын әр глюкоза молекуласынан екі молекула АТФ түзіледі Лимон қышқылы циклінің басқа екі атауы бар: сері Ханс Кребс ашқан Кребс циклы және үшкарбон қышқылдар циклы. Гликолиз бен лимон қышқылы циклының арасындағы басты айырмашылық процестердің клетканың орын алатын жеріне байланысты. Эукариоттарда, гликолиз цитозольде, ал лимон қышқылының циклы митохондрияда орын алады. Лимон қышқылы циклы ферменттерінің көпшілігі митохондрия матриксінде орналасады.
Гликолиз-анаэробтық жағдайда бүлшық етге глюкозаның ыдырауы. Аэробты жағдайда организмде гликолиз лимон қышқылы цикілінің алдында өтеді. Гликолиз процесі нәтижесінде кезектесе жүретін реакция барысында глюкоза пирожүзім қышқылының екі молекуласына ыдырайды. Гликолизге 10 фермент қатысады. Гликолиз процессі клетка цитозолінде жүреді, ол аздаған энергия (8 АТФ) береді. Бүдан өрі қарай пируват митохондрияға енеді, онда лимон қышқылының циклі бойынша аэробтық тотығуға катысады. Бір циклдің жүруі барысында органикалық қышқылдардан төрт сутек (ЗНАДН+ІФАДН) және 2 молекула көмірқышқыл газы CO, бөлініп шығады. Кребс циклі, негізінен, митохондрияның матрикс деп аталатын сұйық фазасында өтеді. Митохондрияда жүретін келесі процесс тотыға фосфорлану деп аталады. НАД және ФАДН кұрамындағы сутек оттектің көмегімен суға дейін тотығады. Бұл процесс кезінде бір молекула НАДН тотықканда 3 молекула АТФ; бір молекула ФАДН тотықканда 2 АТФ синтезделеді. Гликолиз глюкозаның С-6 жағдайы бойынша фосфорланудан басталады да, оныншы реакция кезінде пирожүзім кышкылы мен АТФ түзіледі. Пирожүзім қышқылы аэробты жағдайда СО2 және Н2О-ға дейін толық ыдырайды. БҰЛ кребс циклі, немесе үш карбон қышқылдарының циклі деп аталады.
32.Тыныс алу процессіндегі гликолиздің, анаэробты және аэробты сатыларының ақырғы өнімдері,түзілетін жалпы энергия мөлшері(кай сатыда қанша энергия) есептеңіз. Глюкозаның ыдырауының мысалында энергияның алмасуын қарастырайық. Глюкоза аэробты (О2) және анаэробты (О2-сіз) жағдайларда энергия кљзі бола алады. Дегенмен кљп мљлшердегі энергия органикалық заттардың аэробты ортада ыдырауы кезінде бљлініп шығады. Глюкозаның толық ыдырауы теңдеуінің мысалы: С6 Н12 О6 + 6О2 6СО2 + Н2 О + 2881кДж/моль (3) Глюкоза алдымен плазматикалық мембрана арқылы клетканың цитоплазмасына тасымалданады. Цитоплазманың матриксінде оның оттексіз ыдырауы немесе гликолиз жүреді. Ферменттердің қатысуымен глюкозаның тотыға отырып ыдырауын гликолиз үрдісі дейді.
Гликолиз кљп сатылы үрдіс, оның нәтижесінде глюкозаның бір молекуласынан екі молекула пируват (ПЖҚ – пирожүзім қышқылы) және екі молекула АТФ түзіледі. АТФ-тың синтезделу жолымен танысу үшін гликолиздің сатыларымен танысайық. Гликолиз үш кезеңнен тұрады, атап айтқанда, дайындық кезеңі, гликолиздің оттегісіз және оттегілі кезеңі.
Гликолиздің дайындық кезеңі. Гликолиздің дайындық кезеңі клетканың цитоплазмасында жүреді. Мұнда арнайы ферменттердің әсерінен энергиясы мол органикалық заттар энергиясы аз қарапайым заттарға дейін ыдырайды. Бұл заттар келесі жүретін реакцияладың бастамасы ретінде қолданылады. Химиялық процестерді сызба-нұсқа түрінде былай кљрсетуге болады.
Ақуыз + Н2О SHAPE * MERGEFORMAT аминқышқылдар + Э;
Майлар + Н2Оглицерин + майдың жоғары қышқылы + Э
Полисахаридтер (крахмал, гликоген) + Н2О глюкоза + Э
Гликолиздің оттегісіз (анаәробты) кезеңі.
Бүрдіс митохондриялардың ішкі мембранасында жүреді. Ол ферменттердің әсерімен тірінен кейін бірі кезектесіп жүретін 13 реакцияның жиынтығынан тұрады. реакцияға бастапқы зат ретінде 1 моль С6Н12 О6 (глюкоза) қатысады. Реакция нәтижесінде 2 моль С3 Н6 О3 (сүт қышқылы) және 2 моль АТФ түзіледі.
Бұл реакцияға оттегі мүлде қатыспайды, сондықтан оттегісіз ыдырау деп аталады.
С6 Н12 О6 + 2Н3 РО4 + 2АДФ 2С3 Н6 О3 + 2АТФ + 2Н2 О (4)
Спирттік ашу үрдісі глюкозаның ашуына љте ұқсас кљп сатылы ферменттік ыдырау болып табылады. Оның жиынтық теңдеуі мынадай:
С6 Н12 О6 + 2Н3 РО4 + 2АДФ 2СО2 + 2С2 Н5 ОН+ 2АТФ + 2Н2 О (5)
Яғни, 1 молекула глюкоза ыдырағанда, 2 молекула АТФ синтезеледі. Реакция нәтижесінде энергияның 200 кДж-і немесе 40%-ы АДФ-тың АТФ-қа айналуына жұмсалса, энергияның 120 кДж-і немесе 60%*ы клеткада АТФ түрінде сақталады.
Гликолиздің оттегілік (аэробты) кезеңі. Глюкозаның толық ыдырауы немесе оттегісіз ыдырауы да – митохондрияның ішкі мембранасында үздіксіз жүретін үрдіс. Реакцияға глюкозаның оттегісіз ыдырауы сияқты аралық заттар ретінде ферменттер, АДФ және фосфор қышқылы қатысады. Бұл реакцияның оттегісіз ыдыраудан айырмашылығы – оттегінің қатысуымен глюкоза толық ыдырап, соңғы љнім ретінде СО2 мен Н2 О-ның түзілуі. Реакцияның бастапқы заты ретінде 2 моль С3 Н6 О3 (сүт қышқылы) қатысады. Нәтижесінде 36 моль АТФ синтезделеді. Реакция теңдеуі:
2С3 Н6 О3 + 6О2 + 36Н3 РО4 + 36АДФ 6СО2 + 36АТФ + 42 Н2 О (6)
Яғни, энергияның негізгі кљзі (2600 кДж) глюкозаның оттегілік ыдырауы кезінде түзіледі.
Гликолиздің оттегі ыдырауы кезінде түзілген 26 кДж энергияның 1440 кДж-і немесе 54%-ы АТФ-тың химиялық байланыс энергиясына айналады.
Глюкозаның оттегілік және оттегісіз дырауының жиынтық теңдеуі мынадый:
С3 Н6 О3 + 6О2 + 38Н3 РО4 + 36АДФ 6СО2 + 38АТФ + 44 Н2 О (7)
Глюкозаның оттегілік және оттегісіз ыдырауы кезінде бљлінетін энергияның 80кДж + 1440 кДж = 1520 кДж-і немесе 55%-ы потенциялды энергия түрінде сақталып, клетканың тіршілігіне жұмсалады. Ал 45%-ы жылу энергиясы түрінде пайдаланылады.
Гликолиз процесiне АТФ-тың екi молекуласы пайдаланылған гликолиздегi өнiм 2 АТФ және 2 (НАД*Н + Н+) молекулалары. НАД*Н кейiн өзiнiң сутегi атомын электронтасымалдаушы тiзбекке бередi. Қорыта айтқанда, гликолиз глюкозаны пируватқа айналдырып, НАД*Н және биологиялық қажеттi энергияны АТФ түрiнде түзедi. Гликолиз цитоплазмада жүредi. Гликолизде АТФ-тың таза өнiмi глюкозаның бiр молекуласы АТФ-тың екi молекуласына тең; және лимон қышқылының циклы екi АТФ-ке тең. Митохондрияға химио-осмос теориялық түрде 32 АТФ бере алады. Бiрақ энергияның бiр бөлiгi митохондриядан цитоплазмаға АТФ-тi шығаруға жұмсалуы керек, яғни ол жерде клеткалық АТФ-тiң көп бөлiгi пайдаланылады. Бұл энергияны Н+- резервуарындағы Н+-ионы, және АТФ синтезiне қатыспайтын, АТФ тасымалдауына жұмсалатын иондар түзедi. Сондықтан, митохондрияға химио-осмос нәтижесiнде цитоплазмаға әрбiр глюкоза молекуласынан тек 21 АТФ-ға жуық синтезделiп, бөлiнедi. Әрбiр глюкоза молекуласының ыдырауынан түзiлген АТФ мөлшерiнiң 84%- i ғана Н+- резервуардан келетiн энергия көмегiмен синтезделедi.
Гликолиз,биологиялық маңызы аралық өнімдері мен энергияның түзілуі.Анаэробты ыдырау түрлері,биологиялық маңызы туралы нақты мәлімметтер келтір.
Гликолиз— адам мен жануарлар организмдеріндегі арнайы ферменттердің әсерінен оттегсіз ортада көмірсулардың (негізіненглюкозаның) ыдырау процесі. Жануарлар организміндегі гликолиздің соңғы өнімі —сүтқышқылы. Гликолиз нәтижесінде бөлінгенқуат(энергия) организмнің тіршілігі үшін пайдаланылады. Гликолиз организмдегі тыныс алу және ашу процестерімен тікелей байланысты.Гликолиз - көпсатылы күрделі процесс, ол екі кезеңнен тұрады. Атап айтқанда:1.дайындық кезеңі, 2.АТФ түзелетін кезең.
Гликолиздің дайындық кезеңі жасуша цитоплазмасында жүреді. Мұнда арнайы ферменттердің әсерімен энергиясы мол органикалық заттар энергиясы аз қарапайым заттарға дейін ыдырайды. Мысалы, көмірсулар -моносахаридтерге, майлар -май қышқылы мен глицеринге, нуклеин қышқылдары -нуклеотидтерге, ақуыз - аминқышқылдарына ыдырайды. Бұл заттар келесі жүретін реакциялардың бастамасы ретінде қолданылады. Гликолиз – тынысалу процесi мен көптеген ашу түрлерiне ортақ реакциялардың тiзбегi. Гликолиз кезiнде АТФ-тың аздаған мөлшерi синтезделедi Оттегi жоқ болса, АТФ көзi тек гликолиз болады. Гликолиз реакциясын катализдейтiн ферменттер, цитоплазмада ерiген күйде болады. Сонымен, гликолиз кезiнде глюкозаның алтыбұрышты молекуласы АТФ-тiң төрт молекуласын және екi НАД*Н + Н+ -тi түзе отырып екi пируваттың үшбұрышты молекуласына дейiн ыдырайды.
Гликолиз процесiне АТФ-тың екi молекуласы пайдаланылған гликолиздегi өнiм 2 АТФ және 2 (НАД*Н + Н+) молекулалары. НАД*Н кейiн өзiнiң сутегi атомын электронтасымалдаушы тiзбекке бередi. Қорыта айтқанда, гликолиз глюкозаны пируватқа айналдырып (ол кейiн ары қарай ыдырайды), НАД*Н және биологиялық қажеттi энергияны АТФ түрiнде түзедi. Гликолиз цитоплазмада жүредi. Гликолизде АТФ-тың таза өнiмi глюкозаның бiр молекуласы АТФ-тың екi молекуласына тең; және лимон қышқылының циклы екi АТФ-ке тең. Митохондрияға химио-осмос теориялық түрде 32 АТФ бере алады. Бiрақ энергияның бiр бөлiгi митохондриядан цитоплазмаға АТФ-тi шығаруға жұмсалуы керек, яғни ол жерде клеткалық АТФ-тiң көп бөлiгi пайдаланылады. Бұл энергияны Н+- резервуарындағы Н+-ионы, және АТФ синтезiне қатыспайтын, АТФ тасымалдауына жұмсалатын иондар түзедi. Сондықтан, митохондрияға химио-осмос нәтижесiнде цитоплазмаға әрбiр глюкоза молекуласынан тек 21 АТФ-ға жуық синтезделiп, бөлiнедi. Әрбiр глюкоза молекуласының ыдырауынан түзiлген АТФ мөлшерiнiң 84%- i ғана Н+- резервуардан келетiн энергия көмегiмен синтезделедi.

