




Открытие микроРНК
1993 год. Изучали причины гетерохронных мутаций у нематоды (по Геккелю: макромутации, изменяющие время начала морфогенетического процесса - образование тех или иных органов наступает не во время, что приводит к изменению всего организма). Оказалось, что причина проблем заключалась в том, что некий транскрипционный фактор не исчезал в развитии, хотя должен был. Стали искать ген и нашли мутацию, которая и позволяет этому белку сохраняться дольше.
Lin14 – ядерный белок, фактор транскрипции. В случае мутации lin4, количество белка lin14 не уменьшается. Никакого белка lin4 не кодирует. Lin4 кодирует короткую РНК, комплементарную 3’-концу lin14. Мутацию можно исправить, вводя короткий участок гена.
Тогда не знали про РНК-интерференцию и думали, что это все экзотика, которая свойственна для нематоды. Сейчас мы знаем, что это сложный и свойственный многим организмам процесс, основанный на образовании этих коротких транскриптов.
В 2000 году у нематоды была обнаружена еще одна мутация Let7 (летальна). Этот ген удивительно консервативен, есть у всех многоклеточных, кодирует микроРНК.
2007 год – гетерохронная мутация у кукурузы (изменение дифференцировки меристемы кукурузы). Результат мало похож на кукурузу - много стволов, похож на куст, сначала принимали за сорняки. Сверхэкспрессия двух микроРНК. 2 гена микроРНК, находящиеся рядом, начинают экспрессироваться повышенно → подавление факторов транскрипции. У исследователей создалось впечатление, что образуется новый вид растения: «кукурузная трава».
МикроРНК
22 нт (20-24 н). После процессинга не содержит кэп, 3’-конец не несет 2’-О-СН3-группы (за исключением растений). У siРНК такая модификация есть.
Узнают мишень по механизму РНК-интерференции, несколько сайтов в 3’-НТО. Редкие случаи – узнавание кодирующих районов. Интересно, что есть участки, которые участки узнаются несколькими miRNA, и они будут действовать кооперативно – синергичный эффект
~1000 микроРНК в геномах млекопитающих (4% от общих генов, кодирующих белок), а на самом деле они могут быть модифицированы при синтезе, поэтому реально больше вариантов)
Тканеспецифичная регуляция (на транскрипционном и посттранскрипционном уровне) их образования и функционирования
Кооперация с белковыми транскрипционными факторами (их мРНК – основные мишени для миРНК).
Участие в определении программы развития (стволовые клетки, канцерогенез)
Эволюционный консерватизм (let-7).
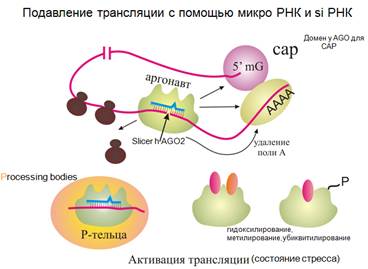
Подавление трансляции с помощью микроРНК
Вы знаете, что мРНК для активной трансляции должна быть в кольцевой форме. Это происходит за счет связанных с кэпом и полиА факторов. Белок Аргонавт (Ago 2) может разрезать соответствующую мРНК, он обладает эндонуклеазной активностью (и при взаимодействии с микроРНК). Меняется конформация струтуры белка и РНК, образуется излом. Чаще это происходит, когда есть совершенная комплементарность (часто наблюдается у растений).
Но очень многие Аргонавты не имеют эндонуклеазной активности. Могут осуществлять (через взаимодействие с другими белками) деаденилирование, диссоциацию рибосом от матрицы, мешать циркуляризации и т.д. У аргонавта есть участок взаимодействия с белком, образующим кэп. Как исключение, Аргонавт иногда может вызывать инициацию трансляции. Кроме того, сам аргонавт может быть модифицирован (гидроксилирование, метилирование,убиквитилирование) и это тоже существенно.
Также, как и с siРНК, Аргонавты имеют положительно заряженную щель, куда ложится миРНК. У человека с миРНК взаимодействует Аргонавт 2, сходный по строению с прочими аргонавтами, даже с аргонавтами термофильных бактерий.
Аргонавт может уходить в цитоплазматические тельца (р-тельца – тельца процессинга). Там происходит деградация РНК, однако Р-тельца также могут служить местами хранения комплекса мРНК, микроРНК и аргонавта, откуда они будут высвобождаться по сигналу. Так можно быстро восстановить подавленную трансляцию.
Аргонавт может узнавать белки, имеющие повторы глицин-триптофан, соединенные гибкими связками, и в этом комплексе идет в Р-тельца
Узнавание мишени
Образуют антипараллельный дуплекс.
Seed – главный детерминант специфичности при выборе мишени, участок, требующий полной комплементарности, 2-8 нуклеотиды на 5’-конце микроРНК. Основа для биоинформатических поисков. Зацепка за мишень начинается с этого seed, а остальные связи дополнительно укрепляют это соединение.
Для расщепления мишени нужна комплементарность нуклеотидов 10 и 11 (мишень на рисунке ниже разрезаться не будет – подавление трансляции без разрезания). Компенсация несовершенства seed иногда осуществляется комплементарностью микро РНК и мишени ближе к 3`-концу микро РНК. Мишени миРНК называют micro RNA targetome.
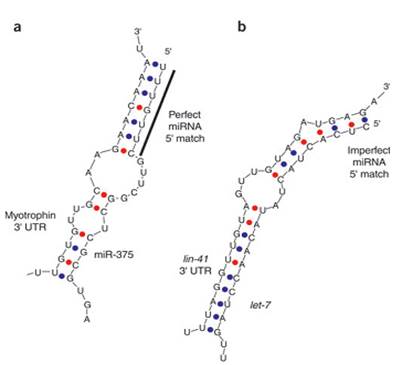
Let7 – важная миРНК и для человека, участвует в клеточной дифференцировке и является антионкогенным фактором.
miRNА вносит вклад в генную экспрессию. Регуляция – подавление может быть разной силы. Тонкая регуляции очень важна для поддержания гомеостаза. Утрата сайта в 3’НТО → освобождение от репрессии → возникновение онкогена. 3’НТО достаточно короткие в генах домашнего хозяйства. Более длинные 3’НТО встречаются в последовательностях матриц регуляторных белков.
Сайты miR очень короткие, легко возникают и исчезают под действием отбора, им приписывается большая роль в эволюции.
Интересен вопрос о биогенезе микроРНК.
Транскрипция предшественника осуществляется Pol II, в редких случаях Pol III (малые ядерные РНК - некодирующие малые РНК, осуществляющие модификацию рРНК – метилирование, введение псевдоуридина. В составе транскриптов этих малых РНК могут быть закодированы предшественники миРНК.). Образуется шпилька-предшественник.
Микропроцессинг осуществляется белком Drosha (РНКаза). На стадии частичного процессинга осуществляется экспорт из ядра в цитоплазму, где он завершается с помощью белка Dicer. Происходит связывание с Аргонавтом.
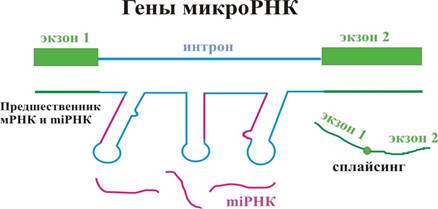
Больше всего миРНК любят находиться в интронах. Несколько микроРНК могут быть закодированы в одном предшественнике. После сплайсинга из шпилечных интронных структур образуются миРНК. Однако у растений иначе: миРНК имеют отдельные гены в межгенных промежутках. Это позволяет говорить о параллельной эволюции системы миРНК у растений и животных.
|
|
На нижнем рисунке нарисован миРНК предшественник.
Где могут находиться транскрипционные единицы (TU) для миРНК? Не только в интронах кодирующих последовательностей (c).
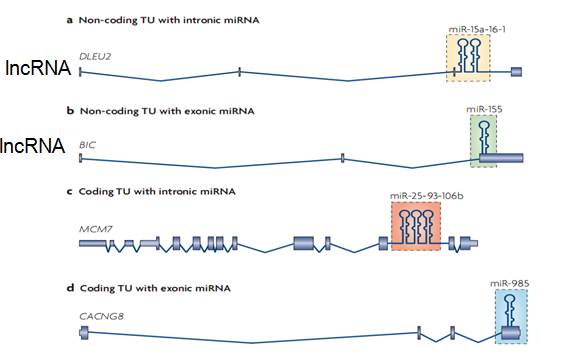
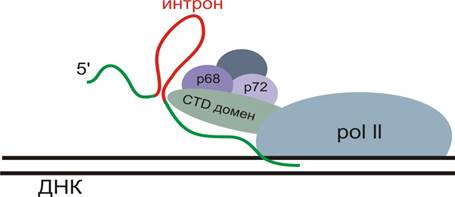
Может находиться в интронах длинных некодирующих РНК (а). Или в экзоне некодирующей РНК (b). Или в экзонах мРНК (d). Сплайсинг мРНК осуществляется котранкрипционно, так же котранскрипционно при участии РНК-хеликаз (РНК-шапероны р68 и р72) будет осуществляться процессинг миРНК. Для этого необходим CTD-домен РНК-полимеразы.
Микропроцессинг
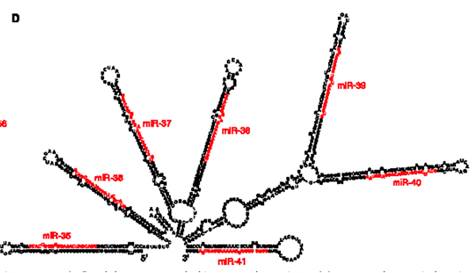
Осуществляет гетеромультимерный комплекс (500-600 кДа). Предшественник миРНК называется Primary, pri-miR, 60-70 н.
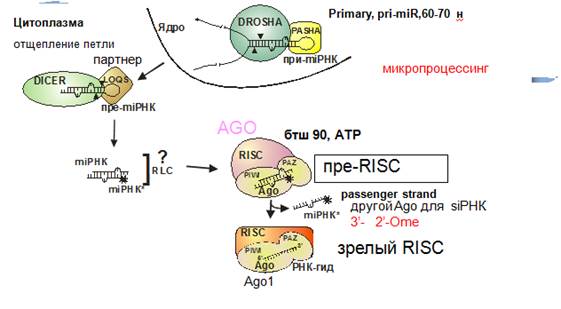 Первой на него садится эндонуклеаза Drosha (узнет дцРНК) имеет 2 домена, нужен Mg, делает 2 разрыва на некотором расстоянии. Образуется структура с терминальной петлей. Сайт расщепления pri-miRNA отмеряется от места перехода одноцепочечной РНК в двуцепочечную. Дальше транскрипт выходит в цитоплазму и Dicer с партнером LOQS осуществляет дальнейшее укорочение в цитоплазме (похоже на siРНК, только партнеры отличаются – пути функционирования и регуляции siРНК и миРНК переплетаюся). Dicer осуществляет расщепление петли, dsРНК попадает в Аргонавт.
Первой на него садится эндонуклеаза Drosha (узнет дцРНК) имеет 2 домена, нужен Mg, делает 2 разрыва на некотором расстоянии. Образуется структура с терминальной петлей. Сайт расщепления pri-miRNA отмеряется от места перехода одноцепочечной РНК в двуцепочечную. Дальше транскрипт выходит в цитоплазму и Dicer с партнером LOQS осуществляет дальнейшее укорочение в цитоплазме (похоже на siРНК, только партнеры отличаются – пути функционирования и регуляции siРНК и миРНК переплетаюся). Dicer осуществляет расщепление петли, dsРНК попадает в Аргонавт.
Способ регуляции:
Если Drosha много, то он атакует шпильку мРНК, которая является матрицей для белка-партнера Drosha → регулируется эффективность.
Та нить, которая будет узнавать мишень – гидовая РНК. miRNA* –комплементарная, passenger strand. Раньше думали, что она уничтожается, однако нашли Аргонавт, который узнает эту нить и использует ее для узнавания некой другой РНК. Для того чтобы miRNA* могла использоваться аргонавтом, она должна быть метилирована на 3’-конце по 2’ОН (так же и у siРНК). Это необходимо, чтобы разные цепи узнавали разные Аргонавты. Чтобы РНК уложилась в аргонавт нужно участие Hsp90 (АТР – зависимый шаперон)→ зрелый RISC.
Обнаружена позитивная регуляция процессинга miR c помощью зрелой miR, комплементарной 3`-концу pri-miR (показано недавно на нематоде) – изменяется конформация предшественника, и он процессируется значительно активнее.

