




Да! Источники:
● взаимно перекрывающиеся комплементарные транскрипты. Впервые обнаружен у растений, затем у животных (защитная и регуляторная функции).
● существование определенных участков в геноме (у дрозофилы в областях, где нет генов – генных пустынях) – палиндромные последовательности. С них получаются т.н. структурные транскрипты → шпильки. (Мишень – мРНК, ответственная за репаративную систему).
● Ооциты млекопитающих. Участвуют ген и псевдоген. Могут быть ориентированы голова к голове (цис-взаимодействовие, в одном месте) или находится в разных участках генома (продукты регулируют за счет транс-взаимодействия). Псевдоген – гомолог гена, который либо плохо транскрибируется (сейчас в статьях их поднимают на уровень генов – они могут даже шпильки образовывать). Псевдоген обычно содержит только экзоны. Образовался путем копирования ДНК на копии мРНК. Псевдогены в ряде случаев несут важную функцию. Псевдоген является супрессором опухоли. Мишенью может быть, например, гистондеацетилаза.
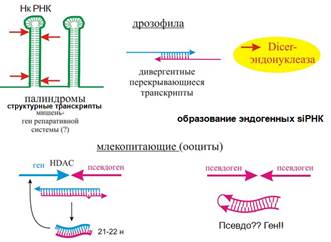
Эндогенные siRNA нашли при изучении косупрессии у растений - оказалось, что если вставить много-много копий трансгена, то можно вообще заткнуть нужный нам трансген (исчезает его эффект), будут растения без пигмента. Назвали косупрессия (cosuppression - homology dependent gene expression), потому что такое ощущение, что трансгены сами себя подавляют.

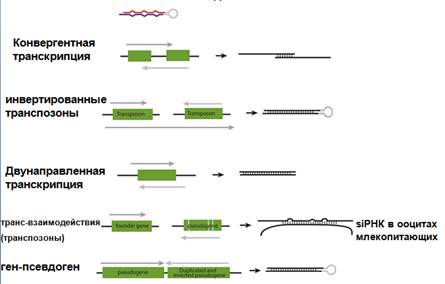
Трансген определял пигментацию лепестков петунии. Дело в том, что трансген вставляется беспорядочно в разной ориентации, и, кроме транскрипта с собственного промотора, есть и транскрипция с других - отсюда перекрывающиеся транскрипты, дцРНК, которых тем больше, чем больше трансгена. Или просто трансген вставился и с него идет транскрипция в обоих направлениях. И так же выяснилось то, чего нет у млекопитающих, но есть у растений и C. elegans - это амплификация siРНК за счет RDRP (РНК-зависимая-РНК-полимераза). Тут короткие РНК могут служить затравками на РНК-мишени, образуются дцРНК, которые снова щепятся Dicer.
У растений в геноме были найдены транскрибируемые участки с обращенными повторами. Такие транскрипты складываются в шпильку и могут служить регуляторами экспресии других генов, давая siРНК. Можно достичь устойчивости к РНК-вирусам растений, если в последнем будет присутствовать гомологичный трансген. Сконструированные вирусы, содержащие последовательности, гомологичные трансгену, неинфекционны.
siРНК используется разными организмами для подавления экспрессии транспозонов (по аналогии с трансгенами растений). PiРНК тоже может выполнять эту функцию, но она есть не во всех тканях организма.
Амплификация siРНК без затравки у растений (например, при косупрессии)
Dicer режет dsРНК, образуется комплекс первично siРНК с аргонавтом, узнаются мишени. Аргонавт привлекает RdRp, но она не использует siРНК как затравку: Аргонавт стимулирует транскрипцию de novo. Аберрантные транскрипты, например, лишенные кэпа, могут являться предпочтительными матрицами для RdRp: косупрессия усиливается при мутации гена 5’-3’ экзонуклеазы XRN4, разрушающей лишенные кэпа мРНК. Образуются короткие вторичные РНК с Р-Р-Р на 5’-конце (это отличает эти siРНК от продуктов нарезки дайсером – там на на 5’-конце, в лучшем случае один фосфат). С ними взаимодействуют вторичные аргонавты, далее эти комплексы могут выполнять самые разнообразные функции в ядре. Можно разрушить эти аберрантные транскрипты. Так амплифицируются РНК, которых много в клетке.
Полученные siРНК, называемые транзитивными, могут передаваться потомству (у нематоды).
У нематоды насчитывают 27 аргонавтов.

И еще раз об источниках эндогенной siРНК
Какие функции выполняет эндо-siРНК у нематоды в ядре.
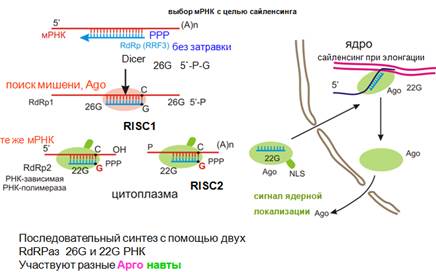
У нематоды образуется небольшое количество коротких РНК, начинающихся с гуанина (такая специфичная РНК-зависимая-РНК-полимераза). Их называют 26G и 22G (присутствует в гораздо большем количестве копий).
В цитоплазме неизвестным образом выбираются мРНК с целью сайленсинга. Этот выбор осуществляется с участием дайсера и RdRP1, действующей без затравки. Образуются монофосфорилированные 26G РНК, которые узнают свою мишень (классическая РНКи). Однако та же мишень может узнаваться другой RdRP2, и здесь синтезируются 22G РНК и привлекается другой Аргонавт (на рисунке он имеет хвостик – сигнал ядерной локализации). В ядре этот комплекс осуществляет сайленсинг при элонгации – узнает насцентный транскрипт. Аргонавт потом можетснова вернуться в цитоплазму. Для некоторых ядерных эндогенных siRNA необходима модификация хвостов, что обеспечит связывание с особым аргонавтом.
Роль 22G (5’-G) siРНК в формировании “центромер” у нематоды (голоцентричные хромосомы).
Напоминаю, что биогенез 22G (5’-G) siРНК идет без участия Dicer
У нематоды нет единого кинетохора, нет центромер, вместо него несколько участков по всей длине. Чтобы эти участки образовывались, должна работать вся эта система - требуется RdRP, терминальная трансфераза (uridylation) (uridylation в отношении микро РНК- повод для ее деградации) и особый Аргонавт (CSR1).
Компоненты этой системы RNAi (Ago, RdRp и нуклеотидилтрансфераза) обнаруживаются в хромосомах, привлекая специфичные центромерные гистоны. Многие Аргонавты функционируют в ядре, связанные с хроматином.

Имеется ген транскрипционного фактора в составе гомеобоксных генов (Hox-кластер) и на некотором расстоянии располагаются регуляторные участки в межгенных промежутках (что-то вроде сайленсеров и энхансеров). На регуляторные участки может садиться белок CTCF (имеет цинковые пальцы, входит в состав белков инсуляторного комплекса). Методом иммунопреципитации в этом комплексе был обнаружен Аргонавт 2, который действует независимо от РНК – белки Аргонавт многофункциональны.

