




Қосспиралді ДНК өлшемдері бір макромолекулаға келетін нуклеотид жұбы (н.ж.) арқылы сипатталады. Клеткалық немесе вирустық ДНК үшін олар үлкен шамада айырмашылығы болады Мысалы,аса көп зерттелген бактериалық плазмидалар көптеген вирус және бактериофаг ДНК-лары бірнеше мың нуклеотид жұбынан (н.ж.) тұрады.Бактериялардың жыныс факторлары, митохондрилар және хлоропласт ДНК-сы бірнеше жүз нуклеотид жұбынан (н.ж.) тұрады. Бактериялардың хромосомдары —бірнеше миллион нуклеотид жұбы (н.ж.), ашытқылапр хромосомдары шамамен 107 нуклеотид жұбын (н.ж.) құрайды.Адам жиынтық хромосомаларының ДНК ұзындығы 3-109 нуклеотид жұбын (н.ж.) құрайды
ДНК-ның нуклеотид кезектілігін анықтайтын заманауи әдістер бір ғана экспериментте 150—300 нуклеотид қалдықтарынан тұратын фрагменттерді секвенирлеуге {ағылш. sequence — кезектілік) мүмкіндік береді.Сондықтан Поэтому ДНК-ның бастапқы молекуласы алдын ала фрагменттеледі.Олүшін көбінесе ДНК-ны рестриктазалармен гидролиздейді.

Бұл фрагменттердің нуклеотид кезектілігін анықтаған соң бүкіл ДНК-ның бірінші реттік құрылымын анықтауға мүмкіндік береді.
ДНҚ-ның нуклеотидтік тізбегін анықтаудың екі принципті айырмашылығы бар әдістері бар.Оның біріншісін А. Максам мен У. Гилберт ұсынған.Ол полинуклеотид тізбекті арнайы химиялық ыдыратуға негізделген. ДНК -ны Максама —Гилберт әдісі бойынша секвенирлегендегі операциялардың реті жалпы түрде мына суретте көрсетілген.
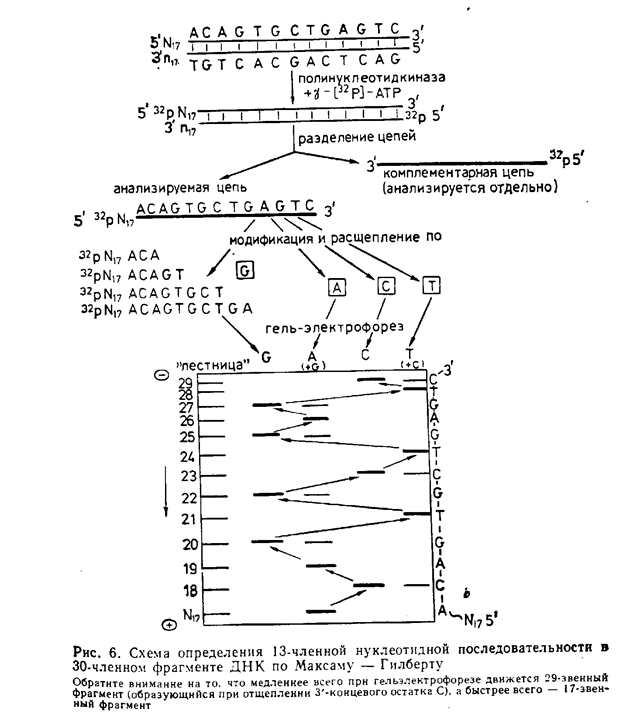
Бұл әдістің негізгі принциптері мынадай:
1. Радиоактивті таңба енгізілген ДНК фрагментінің ұшы гомогенді болуы керек.
2.Анализделетін ДНК тізбегінің үлгісі 4 бірдей порцияға бөлінеді,және химиялық модификация жағдайлары осы 4 порцияның әрқайсысында реакцияға бір немесе екі пурин немесе пиримидин негіздерінің біреуі ғана қатысатындай етіп таңдалады.; пуриндік негіздерді N7-aтом бойынша диметилсульфатпен метилдейді, пиримидинді негіздерді гидразинмен өңдеу арқылы ыдыратады; химиялық реагенттермен өңдеу екі жағдайда да модифицирленген мономерлі звеноның түзілуіне әкеледі,оны полинуклеотид тізбектен бөліп алуға болады.
3. Химиялық модификация шектелген жағдайларда жүргізіледі..
4. Негіздерді бір нуклеотид қалдығна дейін дәлдікпен анықтауға болады.
5 Фрагменттердің ұзындықтарын жоғары сезімтал полиакриламид гелінің жұқа қабатындағы электрофорез әдісі арқылы анықтайды. Электрофорез жоғары температурада 7 М мочевина бар буферлік жүйеде жүргізілгендіктен фрагментердің екінші реттік құрылымы бұзылады
Екінші әдісті Ф. Сэнгер әріптестерімен жасаған.Оның негізінде ДНК-полимеразалық реакция жатыр.Әдістің принципі суретте көрсетілген
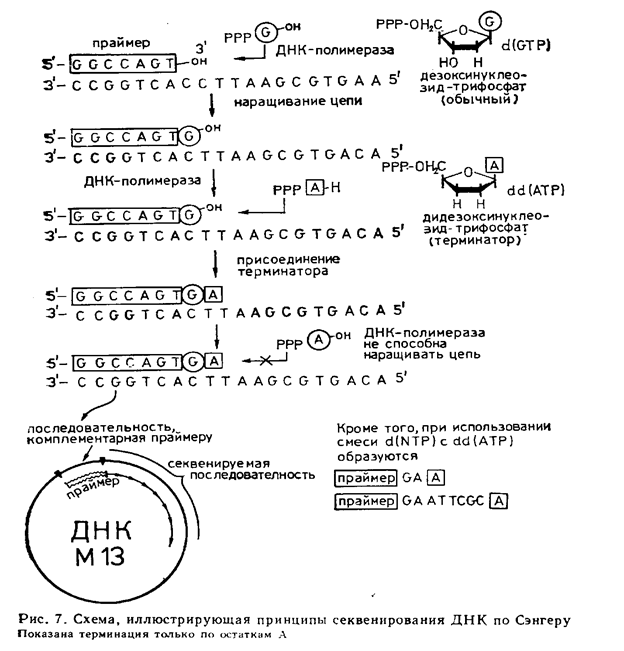
Қазіргі уақытта Максам – Гилберт және Сэнгер әдісі сияқты әр түрлі әдістер бар және оларды толық автоматтандыруға сәті түсті. Дәл осылай, мысалы, ДНК секвенирлеу үшін праймердің 5'- ұшына соңына флоуресценциялық таңбаларды енгізеді, және де әрбір талданатын нуклеотидтардың төртеуінің әртүрлі спектрлік мінездемелерін анықтауда флоуресценциялаушы агенттерді қолданылады. Барлық биохимиялық операциялар роботпен өткізіледі.
ДНК толық нуклеотидті жүйелілігін анықтауда автоматтардың қолдануы принципті түрде кез келген организм геномын адамның геномын да анықтауға мүмкіндік береді.
Лекция 6
ДНҚ репликациясы
Жоспары:
1.ДНҚ репликациясы.Тұқым қуалаудағы ДНҚ-ның ролі.
2.Генетикалық ақпараттың өзін-өзі өндіруі. Матрицалық процестер
3. Репликацияның негізгі принциптері.
4.ДНҚ репликациясының жартылай консервативтілігі механизмінің тәжірибелік дәлелдемесі. Мезельсон жөне Сталь тәжірибесі.
5.Сақиналы ДНҚ репликациясы. Эукариот және прокариот репликациясының ерекшеліктері.
Белок биосинтезінің және нуклейн қышқылдың синтездің басқа биохимиялық реациядан ерекшелігі процеске ферменттпен субстраттан басқа арнайы нуклейн қышқылының молекуласы матрица қатысады. Мұндай процестерді матрицалық процестер деп атайды. Оларға негізінен 3 процесс жатады.
1) Репликация (қайталау).
2) Транскрипция (ағыл: көшіру).
3) Трансляция (аударып көшіру).
Полимер тізбегінің синтезі 3 негізгі элменттен тұрады.
1. Инициация.
2. Элонгация.
3. Терминация.
Инициация деп – түзілетін полимер тізбегінің мономер буындары арасындағы алғашқы байланыстың түзілуін атайды.
Эломгация деп - өсіп бара жатқан полимердің тізбегіне кезекті мономердің қосылуын, яғни тізбектің ұзаруын айтады.
Терминация деп – арнайы сигнал әсерінен тізбектің өсуі мен тоқтауын айтады. Эломгация процестерінің әрбір актісі 3 негізгі элементтен тұрады.
1. Мономердің іріктелуі.
2. Химиялық түрленулер.
3. Транслокация.
Кез келген клетка бөлінер алдында оның ДНҚ молекуласы екі еселенеді және соның нәтижесінде ұрпақ клеткалары алғашқы аналық клеткадағыдай ДНҚ молекуласына ие бо-лады. Олай болса, бөлінетін клетканың ДНҚ-сы дәл өзіне ұқсас тағы бір ДНК молекуласын қалай жасайды?
1940 жылы Л. Полинг пен М. Дельбрюк ген (ДНҚ) өзінше бір бейненің қалыбы секілді, ол қалыпқа саз балшық құйып, оның формасын алуға, содан кейін осы формадан қалып етіп пайдаланған алғашқы форманы қайтадан жасауға болады деген пікір айтқан. Яғни, бұл геннің алғашқы құрылымына комплементарлы ДНҚ құрылымы жасалады, одан алғашқы құрылымға сәйкес ДНК пайда болады деген соз. Шынында да ДНҚ-ның бір тізбегін бір бейне десек, оған комплементарлы екінші тізбек оның кері бейнесі болып табылады. Демек, Уотсон мен Крик көрсеткен ДНК-ның еселенуінің немесе репликациясының жүру жолы шын мәнінде Полинг пен Дельбрюктің болжамын қайталау десе де болғандай.
Сонымен, ДНҚ мынадай жолмен екі еселенеді. Алғаш спиральдың екі тізбегі бір нүктеден бастап ажырай бастайды. Сонан кейін бір-бірінен алшақтап ажыраған әрбір тізбектердің бойына, оларға сәйкес жаңа тізбек синтезделіп, жана тізбек жасалу барысында ажыраған екі тізбекпен өзінің азоттық негіздері арқылы байланысып, онымен өз алдына жаңа спираль құрай бастайды. Сөйтіп алғашқы ДНҚ-ның екі тізбегі толық ажырап болғанда, екі жаңа спи-раль да жасалып бітеді. Алғашқы ДНҚ тізбегі ажырамай тұрғанындағы екінші ескі тізбегіне толық ұқсас боладыӘрине, бұл процесті де клеткадағы ферменттер жүргізедлі. ДНК тізбектерінің бағытгары қарама-қарсы екені белгілі. Жұмысына өте мұқият ферменттер жаңа тізбекті тек бір бағытта, яғни 5'— 3' бағытында ғана жасайды. Олай болса, ферменттер ажыраған тізбектердің біреуінің бойымен жаңа тізбекті жоғарыдан төмен қарай, ал екіншісінің бойымен төменнен жоғары қарай синтездейді. Ең қызығы жаңа тізбектер үздіксіз жасалмайды, ескі тізбектің бойында бірінен кейін бірі шағын ДНҚ фрагменттері пайда болып отырады. Ондай фрагменттердң ұзындығы қарапайым бактерияларда 200 нуклеотидтен тұрса, күрделі организмдерде ол 2000-ға жуык. Осындай фрагменттерді алғаш байқаған жапон ғалымы Р. Оказаки, сондықтан оларды оказаки фрагменттері деп атайды.
1953 жылы Дж. Уотсон жоне Ф. Крик ұсынган ДНК құрылымының үлгісі (моделі) генетикалық хабардың кодын (шартты қысқарту). м у та ц и я л ы қ ө з г е ргіштіктің және гендердін көшірмесінің (ДНК молекуласының бөліктері) а л ы н у ы н т ү с і н у ге мүмкіншілік берді. 1957 жылы М. Мезельсон мен Ф. Сталь, Дж. Уотсоп және Ф. Криктің бактериялық клеткадағы ДНК-ның жартылай консервативті түрде екі еселенуі (репликация) жөніндегі көзқарасын дәлелдеді. Ал Г. Стент ДНҚ екі еселенуінің үш түрін ұсынды:
1) консервативтік (лат. "косервативус" — сақтаушы, негізгі пішіін сақтау) еселенуде ұрпақтық ДНҚ-ларда аналық ДНҚ-нын материалы болмайды;
2) жартылай консервативтілік түрінде ДНҚ-ның жаңа молекуласыиыц бір тізбегі аналық ДНҚ-дан болса, екіншісі — жаңадан құрылған тізбек;
3) дисперсиялык (лат. "дисперсис" — шашырау, бытыраңкы) түрінде аналық ДНҚ-нын материалы кездейсоқ, шашырап жана ДНҚ молекуласында орын алады.
М. Мезельсон мен Ф. Стальдың зерттеулері осы үшеуінің ішінен ДНҚ-ның жартылай консервативті екі еселену түрін таңдап алуға көмектесті. ДНҚ екі еселенуіпің жартылай консервативті жолмен жүруін дәлелдеу Дж. Уотсон мсн Ф. Криктің жасаған ДНҚ молекуласъшың үлгісінің дұрыстығының айғағы болды.
Сонымен ДНҚ еселенуі онын тізбектерінің ажырауынан басталады дедік. Ол тізбекгерді хеликаза (хеликс-спираль) — дезоксирибонуклеаза ферменттсрі — ДНК молекуласының бойымсн екі бағытта жоғары жәнс төмен ажыратады. Нуклеотидтср жұптары меи ДНҚ~ның шиыршықты тізбегінің арасындагы сутектік байланыстары молекуланың бір жақ шетінде бірте-бірте үзіле бастайды және (ДНҚ) тізбектердің екеуі де бірінен бірі босай отырып, жазылады. Осылайша жазылған тізбск, өзінің қосылыстарын оське тік "қоя" отырып, дезоксирибоза және фосфор қышқылының калдықтары арасындағы байланыстар арқылы ұсталып тұрады. Қоршаган ортадан клеткада жинақталған бос нуклеотидтер бар, олар ДНҚ~ ның жазылған тізбегінің бос қосылыстарымен реакцияға түсе алады. Бірақ ол косылысқа бір жұп, "толықтыра түсетін" нуклсотид қана жуықтап, жалғаса алады, Бұл жазылган тізбекке басқа, ДНК,-ның жетіспейтін тізбегі жалғаса бастайды деген сөз. Осы процестердің нәтижесінде ДНҚ-ның екі молекуласы найда болады, олардың әрқайсысында қайтадан жинакталған молекуламен толықтырылған аналық молекуланың жартысы болады. Сонымен түзілген молекулалар ДНК,-ның аналық, молекуласына мейлінше ұксас келсді. Мұнда генетикалык материалдың құрамы да сақталады. Тізбектердің ажырауы мен қосылуы ферменттердің ықпалымен жүреді. Ажыраған тізбектерде оказаки фрагменттері жасала бастайды. Әр фрагмент он шақты нуклеотидтен тұратын РНК, тізбегінен басталады. ДНҚ тізбегінің бойымен РНҚ түріндегі жаңа тізбекті праймаза (РНК, — полимераза) ферменті ғана бастай алады. Тізбекті бастаған РНҚ бөлшегінен ары қарай "ДНҚ — полимераза — 3" деген фермент ажыраған ДНҚ бөлігіне сәйкес етіп оказаки фрагментін синтездейді. Содан кейін басқа "ДНҚ полимераза — 1" ферменті фрагменттердің бастаушысы болған әлгі РНК, тізбегін ыдыратып жібереді. Енді кезек "ДНК-лигаза" деген ферментке келеді, Ол оказаки фрагменттерінің арасын ескі ажыраған тізбекке сәйкес етіп нуклеотидтермен толтырады. Ең соңында "ДНК полимераза - 2" ферменті көптегсн фрагменттердің бірігуінен пайда болған жаңа тізбектің ңуклеотидтерінің ескі тізбегімен сәйкес келетіндігін тексереді. Егер қандай да бір нуклеотид өз орнында тұрмаса соңғы аталған фермент оны кесіп алып тастап, оның орнына тиісті нуклеотидті қояды.
Осындай әр түрлі қызмет аткаратын ферменттердін үйлесімді жұмыс жасауы тұқымдық, белгіні ДНК арқылы-синтездей алады, өиткені матрицалық тізбектің соңы бос 3'— ОН тобынан басталған, дәл осы соңғы бөлікке ғана өсетін ДНҚ тізбегінің келесі нуклеотиді комплементарлық ережеге сәйкес қосыла алады. Бұл тізбектің сннтезделуі, екінші тізбекке қараганда сәл ерте басталады, сол себепті ол «бастаушы» тізбек деп аталады.
ДНҚ-ның екінші 5'—►3' бағытындағы тізбектің («артта қалған») синтезделуі үзілмелі түрде жүреді, Бұл тізбекті ДНҚ-полимераза III ферменті бірден өздігінен синтездей алмайды. «Артта қалган» тізбек синтезін қоздыру үшін ен; алдымен РНҚ-ныц қысқа фрагменті (ұзыпдығы 50 нуклеотидке дейін) қажет. Бұл қысқа кесінді праймер (немесе қоздырыш) деп аталады, олардың синтезделуі праймаза ферменті (РНҚ-полимераза ферментшен өзгсшелігі — иолекулальиқ массасы әлдеқайда төмен және транскрипция процесіне тікелей қатынаспайды) қамтамасыз етеді. Праймердің 3'—• ОН — ұшы ДНҚ-полимераза III ферментіне ДНҚ-ның 5'— >3' бағытындағы тізбегінің синтезделуін бастау үшін қоздырушы болып табылады.

Лекция 7

