




Матричными называются такие процессы, при которых на основе первичной структуры одного биополимера, называемой матрицей, синтезируется первичная структура другого биополимера, называемого копией, причем структура матрицы определяет структуру копии. К матричным процессам относятся:
1.биосинтез ДНК или репликация;
2.биосинтез РНК или транскрипция;
3. биосинтез белка или трансляция.
Любой матричный процесс можно разбить на 3 фазы:
1.начало синтеза или инициация
2.продолжение синтеза или элонгация
3.окончание синтеза или терминация.
Это ферментативные процессы, кроме того, требующие затраты не ферментных белковых факторов. Это энергозависимые процессы, которые требуют затраты энергии в виде АТФ или ГТФ. Ведущим правилом всех матричных процессов является правило комплиментарности. В ходе первых двух процессов, которые в основном осуществляются в ядре клетки, матрицей является нуклеиновые кислоты и копией нуклеиновые кислоты. В процессе транскрибции матрицей является нуклеиновая кислота, а копия полипептидная цепь.
Общая характеристика репликации
Одной из важнейших функций нуклеиновых кислот (НК) является передача генетической информации от родителей потомкам. Этот процесс связан с удвоением, или репликацией, НК (ДНК или РНК), выполняющей функцию хранителя генетической информации, и последующей передачи ее потомкам. Например, в результате деления дочерние клетки получают от материнской идентичные молекулы ДНК, а, следовательно, и идентичную генетическую информацию. При размножении вирусы также передают дочерним вирусным частицам точные копии НК (ДНК или РНК). При половом размножении потомки получают генетическую информацию от обоих родителей. Вот почему дети наследуют признаки обоих родителей. Итак, в результате репликации НК образуются дочерние молекулы, нуклеотидные последовательности которых идентичны как между собой, так и с материнской молекулой НК. Репликации может подвергаться как ДНК, так и РНК (у РНК-содержащих вирусов). В основе репликации лежит принцип комплементарности. Различают три типа репликации: полуконсервативный, консервативный, дисперсный. В случае полуконсервативного типа репликации вновь синтезированная молекула НК состоит из одной материнской и одной дочерней полинуклеотидных цепей. При консервативной репликации вновь синтезированная молекула НК состоит только из дочерних полинуклеотидных последовательностей. При дисперсной репликации вновь синтезированная полинуклеотидная цепь НК состоит из фрагментов дочерних и материнских полинуклеотидных последовательностей. Для прокариот и эукариот характерен полуконсервативный тип репликации. У вирусов встречаются все три типа репликации.
Белки и ферменты, участвующие в репликации ДНК
В репликации ДНК принимают участие многочисленные белки
и ферменты. Рассмотрим их основных представителей.
ДНК-полимеразы
ДНК-полимеразы осуществляют синтез ДНК. Субстратом для
этих ферментов являются дНТФ: дАТФ, дГТФ, дЦТФ и дТТФ. Урав-
нение реакции, катализируемой ДНК-полимеразами, в общем виде
выглядит так:
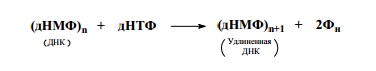
ДНК-полимеразы последовательно добавляют нуклеотиды к
3’ - концу полинуклеотидной цепи:
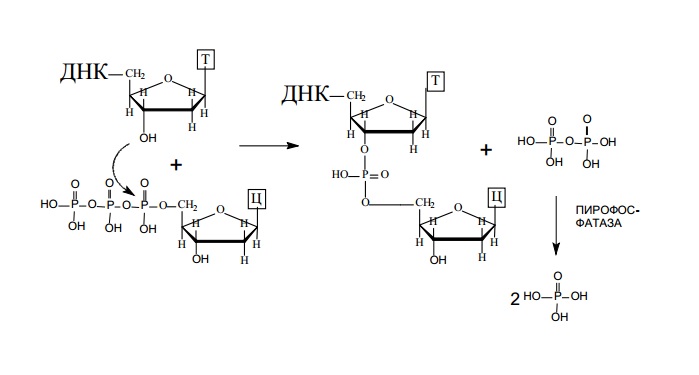
Синтез новой цепи происходит в направлении от 5’-конца к 3’-концу.
ДНК-полимераза может только наращивать цепь ДНК, начать же синтез ДНК с нуля она не в состоянии, для начала ее работы требуется затравка. В качестве затравки может выступать фрагмент ДНК или РНК (рис. 2.1). ДНК-полимераза способна удлинять цепь 23 также только в присутствии цепи, играющей роль матрицы (рис. 2.1). Нуклеотиды присоединяются к затравке в соответствии с принципом комплементарности, напротив аденина всегда будет встраивается
тимин, а напротив гуанина – цитозин (рис. 2.1).

ДНК-полимеразы способны копировать любую цепь ДНК, т.е. они не специфичны к последовательности нуклеотидов матрицы. Скорость ДНК-полимеразной реакции колеблется в пределах от 50 нуклеотидов в секунду у эукариот до 500 нуклеотидов в секунду у прокариот.
Главной задачей ДНК-полимераз является снятие точной копии с матрицы, в связи с этим они проверяют комплементарность каждого нуклеотида дважды: перед включением его в состав растущей цепи и перед тем как включить следующий нуклеотид. Очередная фосфодиэфирная связь образуется, если последний нуклеотид комплементарен матрице. Если включен некомплементарный нуклеотид, то он удаляется за счет 3 -> 5’-экзонуклеазной активности ДНК-полимеразы, и только после удаления некомплементарного нуклеотида ДНК-полимераза продолжит наращивать цепь ДНК (рис.2.2).
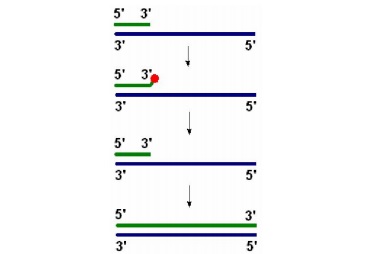
Рис.2.2. ДНК-полимераза удаляет некомплементарный нуклеотид и затем продолжает синтез ДНК.
Праймаза
Праймаза катализирует матричный синтез короткой РНК-
затравки (праймера) в направлении 5’→ 3’. Праймер использует ДНК-полимераза для инициации синтеза ДНК (рис. 2.3).
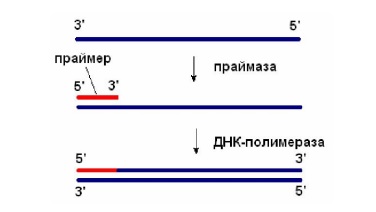
Рис. 2.3. Праймаза синтезирует РНК-затравку, которую ДНК-полимераза использует для синтеза ДНК.
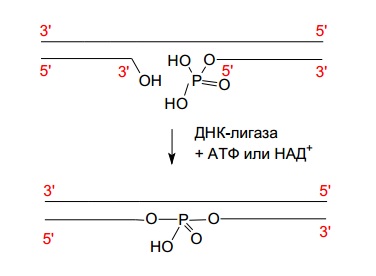
ДНК-лигаза соединяет 5’-фосфатную и 3’-гидроксильную группы соседних нуклеотидов, в результате образуется фосфодиэфирная связь, ликвидирующая разрыв (рис. 2.4).
ДНК-хеликаза осуществляет расплетение двойной спирали ДНК, используя энергию гидролиза АТФ. В результате ее действия возникает «вилка» (Y), состоящая из двуцепочечного участка ДНК и двух одноцепочечных ветвей (рис. 2.5).
Белки, связывающиеся с одноцепочечной ДНК (SSB-белки),
обладают большим сродством к одноцепочечной ДНК, препятствуют
образованию двойной спирали (рис. 2.5).
Топоизомеразы снимают напряжения, возникающее в результате расплетения двойной спирали в ДНК, за счет разрыва и последующего воссоединения цепи ДНК (рис. 2.5).
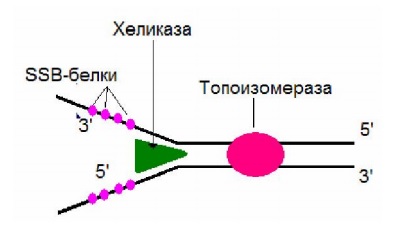
Рис. 2.5. Роль ДНК-хеликазы, SSB-белков, топоизомеразы

