




Пассивный транспорт. Если вещество движется через мембрану из области с высокой концентрацией в сторону низкой концентрации (т. е. по градиенту концентрации этого вещества) без затраты клеткой энергии, то такой транспорт называется пассивным, или диффузией. Различают два типа диффузии: простую и облегченную. Простая диффузия. Характерна для небольших нейтральных молекул (H2O, CO2, O2), а также гидрофобных низкомолекулярных органических веществ. Эти молекулы могут проходить без какого-либо взаимодействия с мембранными белками через поры или каналы мембраны до тех пор, пока будет сохраняться градиент концентрации. Облегченная диффузия. Характерна для гидрофильных молекул, которые переносятся через мембрану также по градиенту концентрации, но с помощью специальных мембранных белков - переносчиков. Для облегченной диффузии, в отличие от простой, характерна высокая избирательность, так как белок переносчик имеет центр связывания комплементарный транспортируемому веществу, и перенос сопровождается конформационными изменениями белка. Один из возможных механизмов облегченной диффузии может быть следующим: транспортный белок (транслоказа) связывает вещество, затем сближается с противоположной стороной мембраны, освобождает это вещество, принимает исходную конформацию и вновь готов выполнять транспортную функцию. Мало известно о том, как осуществляется передвижение самого белка. Другой возможный механизм переноса предполагает участие нескольких белков-переносчиков. В этом случае первоначально связанное соединение само переходит от одного белка к другому, последовательно связываясь то с одним, то с другим белком, пока не окажется на противоположной стороне мембраны.
Митохондрии
Их называют «энергетическими станциями клетки». У эукариот в процессе гликолиза, цикла Кребса и других биохимических реакций формируется большое количество электронов и протонов. Часть из них участвует в разнообразных биохимических реакциях, другая часть аккумулируется в специальных соединениях. Их несколько. Наиболее важные из них НАДН и НАДФН. Эти соединения в форме НАД и НАДФ являются акцепторами – своеобразными «ловушками» электронов и протонов. После присоединения к ним электронов и протонов они превращаются в НАДН и НАДФН и являются уже донорами элементарных частиц. «Отлавливая» их в самых различных частях клетки, они переносят частицы в различные отделы цитоплазмы и, отдавая их на нужды биохимических реакций, обеспечивают бесперебойное течение метаболизма. Эти же соединения поставляют электроны и протоны в митохондрии из цитоплазмы и из матрикса митохондрий, где располагается мощный генератор элементарных частиц – цикл Кребса. НАДН и НАДФН, встраиваясь в цепь переноса электронов, передают частицы на синтез АТФ. Из АТФ энергия черпается на все процессы, идущие в клетке с затратой энергии.
Митохондрии имеют две мембраны жидкостно-мозаичного типа. Между ними располагается межмембранное пространство. Внутренняя мембрана имеет складки – кристы. Внутренняя поверхность крист усеяна грибовидными тельцами, имеющими ножку и головку.
В грибовидных тельцах происходит синтез АТФ. В самой толще внутренней мембраны митохондрий располагаются ферментные комплексы, переносящие электроны с НАДН2 на кислород. Эти комплексы называются дыхательной цепью или цепью пере-

 Рибосома
Рибосома
 А Б С
А Б С
 | |||||||||
 | |||||||||
 | |||||||||
 | |||||||||
 | |||||||||
1 2 3 4 5 6
 Кольцевая ДНК
Кольцевая ДНК
Рис. 16. Митохондрии:
А – Общая схема организации митохондрий. Б – участок кристы с грибовидными телами:
1 – наружная мембрана митохондрий; 2 – межмембранный матрикс; 3 – внутренняя мембрана; 4 – матрикс; 5 – криста; 6 – грибовидные тельца.
носа электронов. За счёт движения э лектронов по этому комплексу происходит синтез АТФ. АТФ является главным поставщиком энергии для всех клеточных процессов. Митохондрии являются главными потребителями кислорода в организме. Поэтому в первую очередь на недостаток кислорода реагируют митохондрии. Реакция эта однозначна –недостаток кислорода (гипоксия) приводит к набуханию митохондрий, в дальнейшем клетки повреждаются и отмирают.
Различные типы эукариотических клеток отличаются друг от друга как по количеству и форме митохондрий, так и по количеству крист. Содержание органелл в клетке колеблется в пределах 500 – 2000, в зависимости от потребности в энергии. Так активно работающие клетки кишечного эпителия содержат много митохондрий, а в сперматозоидах они формируют сеть, обвивающую жгутик, обеспечивая его энергией для движения. В тканях с высоким уровнем окислительных процессов, например в сердечной мышце количество крист во много раз больше, чем в обычных клетках. В митохондриях сердечной мышцы число их в 3 раза больше, чем в митохондриях печени.
Жизнь митохондрий измеряется днями (5 – 20 дней в различных клетках). Устаревшие митохондрии гибнут, распадаются на фрагменты и утилизируются лизосомами. Взамен формируются новые, которые появляются в результате деления имеющихся митохондрий.
Обычно в матриксе митохондрий располагаются 2 – 10 молекул ДНК. Это кольцевые структуры, кодирующие митохондральные белки. В митохондриях имеется весь аппарат синтеза белка (рибосомы, иРНК, тРНК, аминокислоты, ферменты транскрипции и трансляции). Поэтому в митохондриях осуществляются процессы репликации, транскрипции и трансляции, происходит созревание иРНК – процессинг. Исходя из этого, митохондрии являются полуавтономными единицами.
Существенным моментом в деятельности митохондрий является синтез в них стероидных гормонов и некоторых аминокислот (глутаминовой). Устаревшие митохондрии могут выполнять депонирующую функцию – накапливать продукты экскреции или аккумулировать вредные вещества, попавшие в клетку. Понятно, что в этих случаях митохондрия перестаёт выполнять свою основную функцию.
Функции митохондрий:
· накопление энергии в форме АТФ,
· депонирующая,
· синтетическая (синтез белков, гормонов, аминокислот).
Рибосомы
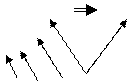
Это аппарат синтеза белка в клетке. В рибосому входят две субъединицы – большая и малая. Субъединицы имеют сложную конфигурацию (см. рис. 14) и состоят из белков и рибосомальной РНК (рРНК). Рибосомальная РНК служит своеобразным каркасом, на который крепятся молекулы белка.
Образование рибосом происходит в ядрышке ядра клетки. Сформированные большая и малая субъединица выходят через ядерные поры в цитоплазму.
В цитоплазме рибосомы находятся в диссоциированном или диспергированном состоянии, это диссоциированные рибосомы. В таком состоянии они не способны прикрепиться к мембране. Это не рабочее состояние рибосомы. В рабочем состоянии рибосома представляет собой органоид, состоящий из двух скрепленных между собой субъединиц, между которыми проходит нить иРНК. Такие рибосомы могут свободно «плавать» в цитозоле, они называются свободные рибосомы, или прикрепляться к различным мембранам,
 |

А Б В Г
Рис. 14. Естественная форма малой (А) и большой (Б) субъединицы рибосомы. Целая рибосома (В). Схематическое изображение рибосомы (Г)
например к мембране ЭПС. На мембране рибосома чаще всего располагается не в одиночку, а ансамблем. В ансамбле может быть разное количество рибосом, но все они соединены одной нитью иРНК. Это делает работу рибосом очень эффективной. В то время как очередная рибосома заканчивает синтез белка и сходит с иРНК, другие этот синтез продолжают, находясь в различных местах молекулы РНК. Ансамбль таких рибосом называется полисомой). Обычно на мембранах шероховатой ЭПС синтезируются белки на экспорт, а в гиалоплазме – на нужды клетки. Если при заболевании обнаруживается отсоединение рибосом от мембран и переход их в гиалоплазму, то это можно рассматривать как защитную реакцию – с одной стороны, клетки сокращают экспорт белка и увеличивает синтез белка на внутренние нужды. С другой стороны, такое отсоединение рибосом свидетельствует о наступающем энергодифеците клетки, так как прикрепление и удержание рибосом на мембранах требует затраты энергии, основным поставщиком которой в клетке является АТФ. Недостаток АТФ закономерно приводит не только к отсоединению рибосом от мембраны, но и неспособности свободных рибосом прикрепиться к мембране. Это приводит к выключению из молекулярного хозяйства клетки эффективного генератора белка – шероховатой ЭПС. Считается, что энергодефицит – это серьёзное нарушение клеточного метаболизма, чаще всего связанное с нарушением в деятельности энергозависимых процессов (например в митохондриях).
В рибосоме имеются три различных участка, с которыми связывается РНК - один для матричной, или информационной РНК (мРНК, или иРНК), и два для транспортной РНК. Первый располагается в месте контакта большой и малой субъединицы. Из двух последних - один участок удерживает молекулу тРНК и формирует связи между аминокислотами (пептидные связи), поэтому его называют Р-центр. Он располагается в малой субъединице. А второй служит для удержания только что прибывшей молекулы тРНК, нагруженной аминокислотой. Его называют А-центром.
Следует подчеркнуть, что при синтезе белка некоторые антибиотики могут блокировать этот процесс.
Эндоплазматическая сеть
Это система каналов и полостей, стенки которых состоят из одного слоя мембраны. Строение мембраны аналогично плазмалемме (жидкостно-мозаичное), однако входящие сюда липиды и белки несколько отличаются по химической организации. Различают два типа ЭПС: шероховатую (гранулярная) и гладкую (агранулярная).
ЭПС обладает несколькими функциями.
- Транспортная.
- Мембранообразующая.
- Синтезирует белок, жиры, углеводы и стероидные гормоны.
- Обезвреживает токсические вещества.
- Депонирует кальций.
На внешней поверхности мембраны шероховатой ЭПС происходит синтез белка.
2. На мембране гладкой ЭПС располагаются ферменты которые синтезируют жиры, углеводы и стероидные гормоны.
3. На мембране гладкой ЭПС располагаются ферменты, которые обезвреживают токсические чужеродные вещества попавшие в клетку.
Шероховатая содержит на внешней стороне матрикса мембраны большое число рибосом, которые участвуют в синтезе белка. Синтезированный на рибосоме белок через специальный канал попадает в полость ЭПС (рис. 7) и оттуда разносится в различные части цитоплазмы (в основном он попадает в комплекс Гольджи). Это характерно для белков, идущих на экспорт. Например, для пищеварительных ферментов, синтезируемых в клетках поджелудочной железы.
 Рибосома иРНК
Рибосома иРНК
А Б В
Рис. 7. Эндоплазматическая сеть:
А – фрагменты гладкой ЭПС; Б – фрагменты шероховатой ЭПС. В – функционирующая рибосома на шероховатой ЭПС.
В мембране гладкой ЭПС имеется набор ферментов, синтезирующих жиры и простые углеводы, а также стероидные гормоны, необходимые для организма. Особо следует отметить, что в мембране гладкой ЭПС клеток печени располагается система ферментов, осуществляющих расщепление чужеродных веществ (ксенобиотиков), попавших в клетку, в том числе и лекарственных соединений. Система состоит из разнообразных белков-ферментов (окислителей, восстановителей, ацетиляторов и др.).
Ксенобиотик или лекарственное вещество (ЛВ), взаимодействуя последовательно с определёнными ферментами, изменяет свою химическую структуру. В результате конечный продукт может сохранить свою специфическую активность, может стать неактивным или, наоборот, приобрести новое свойство – стать токсичным для организма. Система ферментов, расположенная в ЭПС и осуществляющая химическое преобразование ксенобиотиков (или ЛВ), носит название система биотрансформации. В настоящее время этой системе придают большое значение, т.к. от интенсивности её работы и количественного содержания в ней тех или иных ферментов зависит специфическая активность ЛВ (бактерицидность и т.п.) в организме и их токсичность. 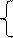
Изучая содержание в крови противотуберкулёзного вещества изониазида, исследователи столкнулись с неожиданным явлением. При приёме одинаковой дозы препарата его концентрация в плазме крови у разных индивидуумов оказалась неодинаковой. Выяснилось, что у людей с интенсивным процессом биотрансформации изониазид быстро ацетилируется, превращаясь в другое соединение. Поэтому его содержание в крови становится значительно меньше, чем у индивидуумов с низкой интенсивностью ацетилирования. Логично сделать вывод, что пациентам с быстрым ацетилированием, для эффективного лечения, необходимо назначать более высокие дозы препарата. Однако возникает другая опасность, при ацетилировании изониазида образуются токсичные для печени соединения. Поэтому повышение дозы изониазида у быстрых ацетиляторов может обернуться поражением печени. Вот такие парадоксы постоянно встречаются на пути фармакологов при изучении механизма действия препаратов и систем биотрансформации. Поэтому один из важных вопросов, которые должен решить фармаколог – рекомендовать для внедрения в практику такое ЛВ, которое не подвергалось бы быстрому инактивированию в системе биотрансформации и, тем более, не превращалось бы в токсическое для организма соединение. Известно, что из рекомендованных в настоящее время Фармкомитетом ЛВ практически все подвергаются процессам биотрансформации. Однако ни одно из них полностью не теряет своей специфической активности и не наносит существенного вреда организму. Такие вещества, как атропин, левомицетин, преднизолон, норадреналин и множество других полностью сохраняют свои свойства, но проходя через систему биотрансформации, становятся более растворимы в воде. Значит, они достаточно быстро будут выводиться из организма. Есть вещества, которые активируют систему биотрансформации, например, фенобарбитал. Так, в проводимых на мышах экспериментах выяснили, что при попадании в кровоток большого количества этого вещества в клетках печени поверхность гладкой ЭПС за несколько дней удваивается. Стимуляцию системы биотрансформации используют для нейтрализации токсических соединений в организме. Так, фенобарбитал используется при лечении гемолитической болезни новорождённых, когда стимуляция систем биотрансформации помогает организму справиться с избытком вредных веществ, например, билирубином. Кстати, после удаления вредного вещества избыток мембран гладкой ЭПС разрушается с помощью лизосом, и через 5 дней сеть приобретает нормальный объем.
Синтезированные в мембранах ЭПС вещества по каналам доставляются к различным органоидам или в места, где они необходимы. Транспортная роль ЭПС этим не ограничивается, в некоторых участках мембрана способна образовывать выпячивания, которые перешнуровываются и отрываются от мембраны, формируя пузырёк, в котором содержатся все ингредиенты канальца сети. Этот пузырёк способен перемещаться и опорожнять своё содержимое в самых различных местах клетки, в частности сливаться с комплексом Гольджи. Необходимо отметить важную роль ЭПС в строительстве всех внутриклеточных мембран. Здесь начинается самый первый этап такого строительства.
Существенную роль играет ЭПС и в обмене ионов кальция. Этот ион имеет большое значение в регуляции клеточного метаболизма, изменяя проницаемость мембранных каналов, активируя различные соединения в цитоплазме и т.д. Гладкая ЭПС является депо ионов кальция. При необходимости кальций высвобождается и принимает участие в жизнедеятельности клетки. Эта функция более всего свойственна ЭПС мышц. Освобождение ионов кальция из ЭПС является звеном в сложном процессе сокращения мышцы.
Необходимо отметить тесную связь ЭПС с митохондриями - энергетическими станциями клетки. При заболеваниях, связанных с энергодифецитом, рибосомы отсоединяются от мембраны шероховатой ЭПС. Последствия не трудно предсказать – нарушается синтез белков на экспорт. А поскольку к таким белкам относятся пищеварительные ферменты, то при заболеваниях, связанных с энергодифецитом, будет нарушена работа пищеварительных желёз и, как следствие, пострадает одна из основных функций организма – пищеварительная. Исходя из этого, должна вырабатываться и фармакологическая тактика врача.
Аппарат Гольджи
В железах внутренней секреции, например, в поджелудочной железе, некоторые пузырьки, отделяясь от ЭПС, уплощаются, сливаются с другими пузырьками, накладываются друг на друга, как блины в стопке, образуя комплекс Гольджи (КГ). Состоит он из нескольких структурных элементов – цистерн, пузырьков и трубочек (рис. 9). Все эти элементы образованы однослойной мембраной жидкостно-мозаичного типа. В цистернах происходит «созревание» содержимого пузырьков. Последние отшнуровываются от комплекса и передвигаются в цитозоле по микротрубочкам, фибриллам и филаментам. Однако основной путь пузырьков – движение к плазматической мембране. Сливаясь с ней, пузырьки опорожняют своё содержимое с пищеварительными ферментами в межклеточное пространство. Из него ферменты попадают в проток и изливаются в кишечник. Процесс выведения при помощи пузырьков секрета КГ носит название экзоцитоз.
 1
1
 2
2
 ЭПС
ЭПС

3 4 5
Рис. 9. Срез комплекса Гольджи: 1 – ядро; 2 – ядрышко; 3 – пузырьки, образующиеся в КГ; 4 – цистерны КГ; 5 – трубочка.
Следует отметить, что экзоцитоз в клетке часто совмещён с другим важным клеточным процессом – строительством или обновлением плазматической мембраны. Суть его в том, что пузырёк, состоящий из однослойной жидкостно-мозаичной мембраны, подойдя к мембране, разрывается, разрывая одновременно и мембрану. После выхода содержимого пузырька его края сливаются с краями бреши в мембране, и разрыв «затягивается». Другой путь характерен для пузырьков, из которых в дальнейшем формируются лизосомы. Эти пузырьки, перемещаясь по направляющим филаментам, распределяются по всей цитоплазме клетки.
Практически в КГ происходит перераспределение белков, синтезированных на рибосомах шероховатой ЭПС и доставленной по каналам ЭПС в КГ, часть из них идёт из КГ на экспорт, часть остаётся для нужд клетки (например, концентрируется в лизосомах). Процесс точного распределения белков имеет сложный механизм, и при его сбоях могут пострадать не только функции пищеварения, но и все функции, связанные с лизосомами. Некоторые авторы очень точно подметили, что КГ в клетке является «центральным железнодорожным вокзалом», где происходит перераспределение потока пассажиров-белков.
Некоторые микротрубочки слепо заканчиваются.
В КГ осуществляется модификация продуктов, поступающих из ЭПС:
1. Накопление поступающих продуктов.
2. Обезвоживание их.
3. Необходимая химическая перестройка (созревание).
Ранее мы отмечали, что в КГ происходит формирование пищеварительных секретов и лизосом. Кроме этих функций, в органоиде синтезируются полисахариды и одни из основных участников иммунных реакций в организме - иммуноглобулины.
И, наконец, КГ принимает активное участие в построении и обновлении плазматической мембран. Изливаясь через плазмалемму, пузырьки способны интегрировать в неё свою мембрану. Для строительства мембран используются вещества, синтезированные в ЭПС и "созревшие" на мембранах цистерн КГ.
Функция КГ:
· транспортная (образовавшиеся пузырьки транспортируют ферменты наружу или для собственного использования),
· формирует лизосомы,
· образующая (в КГ образуются иммуноглобулины, сложные сахара, мукопротеиды и т.д.),
· строительная: а) мембрана пузырьков КГ может встраиваться в плазматическую мембрану; б) на строительство мембран клетки идут соединения, синтезированные в мембране цистерн,
· разделительную (делит клетку на отсеки).

