




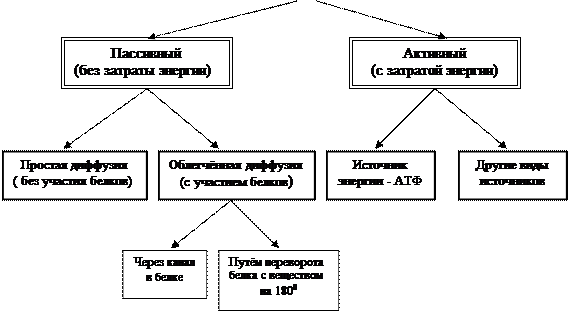
· Перенос веществ через мембрану. Через мембрану осуществляется транспорт веществ в обе стороны мембраны.
· Перенос информации через мембрану. На мембране информация из вне воспринимается преобразуется и передаётся в клетку или из клетки. Существенную роль при этом играют рецепторы мембран.
· Защитная роль. а) защищает содержимое клетки от механических повреждений, химических реагентов и биологической агрессии, например от проникновения вирусов и др.;
б) в многоклеточном организме рецепторы плазматической мембраны формируют иммунный статус организма;
в) в многоклеточном организме мембрана обеспечивает протекание реакции фагоцитоза.
- Ферментативная - в мембранах находятся различные ферменты (например, фосфолипаза А и др.), которые осуществляют целый ряд ферментативных реакций.
- Гликопротеины и гликолипиды на цитоплазматической мембране осуществляют контакт с мембранами других клеток.
 | |||
 |
Цитоскелет
Цитоскелет — это клеточный каркас или скелет, находящийся в цитоплазме живой клетки. Он присутствует во всех клетках как у эукариот, так и у прокариот. Это динамичная, изменяющаяся структура, в функции которой входит поддержание и адаптация формы клетки ко внешним воздействиям, экзо- и эндоцитоз, обеспечение движения клетки как целого, активный внутриклеточный транспорт и клеточное деление. Цитоскелет образован белками. В цитоскелете выделяют несколько основных систем, называемых либо по основным структурным элементам, заметным при электронно-микроскопических исследованиях (микрофиламенты, промежуточные филаменты, микротрубочки), либо по основным белкам, входящим в их состав (актин-миозиновая система, кератины, тубулин-динеиновая система). Цитоскелет эукариот. Клетки эукариот содержат три типа так называемых филаментов. Это супрамолекулярные, протяжённые структуры, состоящие из белков одного типа, сходные с полимерами. Разница заключается в том, что в полимерах связь между мономерами ковалентная, а в филаментах связь составных единиц обеспечивается за счёт слабого нековалентного взаимодействия. Актиновые филаменты (микрофиламенты)Порядка 7 нм в диаметре, микрофиламенты представляют собой две цепочки из мономеров актина, закрученные спиралью. В основном они сконцентрированы у внешней мембраны клетки, так как отвечают за форму клетки и способны образовывать выступы на поверхности клетки (псевдоподии и микроворсинки). Также они участвуют в межклеточном взаимодействии (образовании адгезивных контактов), передаче сигналов и, вместе с миозином — в мышечном сокращении. С помощью цитоплазматических миозинов по микрофиламентам может осуществляться везикулярный транспорт. Промежуточные филаменты. Диаметр промежуточных филаментов составляет от 8 до 11 нанометров. Они состоят из разного рода субъединиц и являются наименее динамичной частью цитоскелета. Микротрубочки. Микротрубочки представляют собой полые цилиндры порядка 25 нм диаметром, стенки которых составлены из 13 протофиламентов, каждый из которых представляет линейный полимер из димера белка тубулина. Димер состоит из двух субъединиц — альфа- и бета- формы тубулина. Микротрубочки — крайне динамичные структуры, потребляющие ГТФ в процессе полимеризации. Они играют ключевую роль во внутриклеточном транспорте (служат «рельсами», по которым перемещаются молекулярные моторы — кинезин и динеин), образуют основу аксонемы ундилиподий и веретено деления при митозе и мейозе.
Эукариотические хромосомы
Хромосомы — нуклеопротеидные структуры в ядре эукариотической клетки (клетки, содержащей ядро), которые становятся легко заметными в определённых фазах клеточного цикла (во время митоза или мейоза). Хромосомы представляют собой высокую степень конденсации хроматина, постоянно присутствующего в клеточном ядре. В хромосомах сосредоточена большая часть наследственной информации. Основу хромосомы составляет линейная (не замкнутая в кольцо) макромолекула дезоксирибонуклеиновой кислоты (ДНК) значительной длины (например, в молекулах ДНК хромосом человека насчитывается от 50 до 245 миллионов пар азотистых оснований). В растянутом виде длина хромосомы человека может достигать 5 см. Помимо неё, в состав хромосомы входят пять специализированных белков (так называемые гистоны) и ряд негистоновых белков. Последовательность аминокислот гистонов высококонсервативна и практически не различается в самых разных группах организмов.
В интерфазе хроматин не конденсирован, но и в это время его нити представляют собой комплекс из ДНК и белков. Макромолекула ДНК обвивает октомеры (структуры, состоящие из восьми белковых глобул) гистоновых белков H2A, H2B, H3 и H4, образуя структуры, названные нуклеосомами. В целом вся конструкция несколько напоминает бусы. Последовательность из таких нуклеосом, соединённых белком H1, называется нуклеофиламентом, или нуклеосомной нитью, диаметром около 10 нм.
В ранней интерфазе (фаза G1) основу каждой из будущих хромосом составляет одна молекула ДНК. В фазе синтеза (S) молекулы ДНК вступают в процесс репликации и удваиваются. В поздней интерфазе (фаза G2) основа каждой из хромосом состоит из двух идентичных молекул ДНК, образовавшихся в результате репликации и соединённых между собой в районе центромерной последовательности.
Перед началом деления клеточного ядра хромосома, представленная на этот момент цепочкой нуклеосом, начинает спирализовываться, или упаковываться, образуя при помощи белка H1 более толстую хроматиновую нить, или хроматиду, диаметром 30 нм. В результате дальнейшей спирализации диаметр хроматиды достигает ко времени метафазы 700 нм. Значительная толщина хромосомы (диаметр 1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой микроскоп. Конденсированная хромосома имеет вид буквы X (часто с неравными плечами), поскольку две хроматиды, возникшие в результате репликации, по-прежнему соединены между собой в районе центромеры (подробнее о судьбе хромосом при клеточном делении см. статьи митоз и мейоз).
Первичная перетяжка
Хромосомная перетяжка (X. п.), в которой локализуется центромера и которая делит хромосому на плечи.
Вторичные перетяжки
Морфологический признак, позволяющий идентифицировать отдельные хромосомы в наборе. От первичной перетяжки отличаются отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают короткими и длинными и локализуются в разных точках по длине хромосомы. У человека это 9, 13, 14, 15, 21 и 22 хромосомы.
Типы строения хромосом
Различают четыре типа строения хромосом:
- телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце);
- акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом);
- субметацентрические (с плечами неравной длины, напоминающие по форме букву L);
- метацентрические (V-образные хромосомы, обладающие плечами равной длины).
Тип хромосом является постоянным для каждой гомологичной хромосомы и может быть постоянным у всех представителей одного вида или рода.
Такие хромосомы, для которых характерны огромные размеры, можно наблюдать в некоторых клетках на определённых стадиях клеточного цикла. Например, они обнаруживаются в клетках некоторых тканей личинок двукрылых насекомых (политенные хромосомы) и в ооцитах различных позвоночных и беспозвоночных (хромосомы типа ламповых щёток). Именно на препаратах гигантских хромосом удалось выявить признаки активности генов.
Политенные хромосомы
Впервые обнаружены Бальбиани в 1881-го, однако их цитогенетическая роль была выявлена Костовым, Пайнтером, Гейтцем и Бауером. Содержатся в клетках слюнных желёз, кишечника, трахей, жирового тела и мальпигиевых сосудов личинок двукрылых.
Хромосомы типа ламповых щёток — это специальная форма хромосом, которую они приобретают у многих животных в ходе оогенеза (развития женских половых клеток). Впервые обнаружены Флеммингом в 1882 году; само название «хромосомы типа ламповых щёток» было предложено Рюккертом в 1892 году. По длине превышают политенные хромосомы, наблюдаются в ооцитах на стадии диплотены первой профазы мейоза, во время которой процессы синтеза, приводящие к образованию желтка, наиболее интенсивны. Общая длина хромосомного набора в ооцитах некоторых хвостатых амфибий достигает 5900 мкм.

