




Эффективность осеменения зависит не только от спермия, но и от судьбы яйцеклетки после ее выделения из фолликула. В момент овуляции яйцевая клетка, окруженная лучистой короной (клетки фолликулярного эпителия), вместе с фолликулярной жидкостью выделяется в брюшную полость (рис. 39).
Вследствие тесной связи яйцепровода с яичником яйцевая клетка обычно попадает на ответвления бахромки. Этому же способствуют помимо анатомических условий динамические свойства бахромки и яйцепровода. Так, у обезьян к моменту овуляции сокращаются мышцы яйцепровода и маточной брыжейки.
Бахромка вследствие наполнения ее сосудов кровью становится упругой и охватывает яичник, как бы подготавливая ложе для выделяющегося яйца.
У кобылы, свиньи, собаки, самки медведя, хорька и у некоторых других животных яичник заключен в бахромку, или складку, яйцепровода; таким образом создаются такие взаимоотношения, при которых выделившееся яйцо, как правило, должно попасть непосредственно на бахромку. У других животных (корова, овца, коза) бахромка развита сравнительно слабо и не может охватить овуляционный участок яичника, поэтому нельзя объяснить проникновение яйца в яйцепровод исключительно эректильным состоянием бахромки. Процесс этот некоторые авторы объясняют токами жидкости из брюшной полости в матку. Возбудителем этих токов В. С. Груздев считает колебания ресничек эпителия бахромки и яйцепрово- да. Мерцательные движения их, направленные в полость матки, перегоняют жидкость из брюшной полости в матку, а образовавшиеся токи увлекают выпавшую в брюшную полость яйцевую клетку. Такого же мнения придерживается К. К. Скробанский, который объясняет возникновение токов жидкости еще и сокращением брюшного пресса, повышающим внутрибрюшное давление в момент овуляции.
По наблюдениям медиков, в большинстве случаев трубной беременности яйцевая клетка выделяется правым яичником, а плод развивается в левой трубе. Это обстоятельство позволило предположить, что после овуляции яйцевая клетка может «странствовать» по брюшной полости, а затем попадать в тот или иной яйцепровод. Специальные опыты послужили основанием к введению понятий о наружном и внутреннем «странствовании» яйца или зиготы (тщга1ю ех(;егпа е1 гт§га(:ю т(:егпа). Первое заключается в том, что яйцевая клетка по выделении ее из яичника некоторое время плавает в капиллярных щелях между петлями кишечника и лишь позднее проникает в полость матки через просвет одной из труб. Внутреннее «странствование» выражается в перемещении зиготы (возможно, и зародыша) из одного рога в другой через тело матки и может быть объяснено сокращением мышцы матки.
Сроки продвижения яйцевых клеток по яйцепроводам, по-видимому, могут сильно варьировать. Специальные опыты и клинические наблюдения позволяют допустить, что прохождение яиц по яйцепроводам у всех животных продолжается 1—3 сут. У коров яйцо в течение
 5
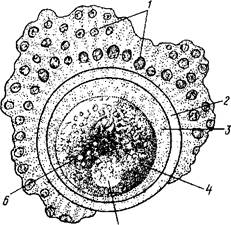
Рис. 39. Схема строения яйца:
5
Рис. 39. Схема строения яйца:
|
1 — клетки лучистого венца; 2— прозрачная оболочка; 3— перивителлиновое пространство; 4— цитоплазма; 5 — ядро; 6— желточные зерна
первых 6—12 ч после овуляции проходит треть яйцепровода. Дальнейшее продвижение яйцеклетки происходит медленно — примерно около 0,1см в 1 ч. После овуляции яйцеклетка способна к оплодотворению в течение 4—6 ч.
У птиц этот срок составляет всего только 20—25 мин (А. Д. Курбатов и др.). Если оплодотворения не произошло, яйцо погибает и рассасывается. Время жизни яйцеклеток у некоторых животных и поступление зигот в матку приведены в таблице 12.
12. Сроки жизни яйцевых клеток, продвижения зигот и имплантации зародышей (по Р. X. Хантеру)
|
Оплодотворение — физиологический процесс, заключающийся в слиянии яйца и спермиев с последующей их ассимиляцией и диссимиляцией, в результате чего образуется новая клетка (зигота), обладающая двойной наследственностью.
В 1651 г. У. Гарвей высказал свое историческое изречение: «Все новое — из яйца», породившее направление овистов (оуо — яйцо).
После открытия спермиев студентом Гаммом и его учителем А. Г. Левенгуком (1677 г.) в биологии стала господствовать теория преформизма, утверждавшая, что спермий, являясь микроскопически малым, но вполне сформированным организмом (атта!си1ит — зверек, Ьотип- си1ит — человек), попадает во время полового акта в полость матки и использует материнский организм как почву для своего роста (не развития!). Поэтому в русской литературе еще встречаются два порочных термина для обозначения спермиев: сперматозоид (семя, подобное животному) и живчик (живой человечек).
В 1759 г. русский академик К. Ф. Вольф опубликовал свой труд «Теория зарождения», которым положил начало теории эпигенеза, т. е. теории развития. С этого времени началась двухвековая борьба сторонников двух течений: эпигенеза и преформизма.
В 1827 г. русский ученый К. Бэр открыл яйцевую клетку млекопитающих и заложил основы современной эмбриологии. После открытия яйцеклетки одни преформисты усматривали готовый микроорганизм в спермии, другие — в яйце.
В 1876 г. Р. Гертвиг и Фоль впервые наблюдали проникновение спермия в яйцевую клетку морского ежа. Наука стала приближаться к пониманию сущности оплодотворения.
Во время оплодотворения происходят взаимная ассимиляция (а881тйа1ю — взаимопоглощение) и диссимиляция (ё1881гш1а1ю — расподобление) яйца и спермия, в результате которых уже не существует ни яйца, ни спермия, а образуется новая, третья клетка — зигота (от греч. 2у§о1о8 — соединенный вместе), не тождественная ее двум первоисточникам. Поэтому широко применяемое в биологии называние зиготы оплодотворенным яйцом не соответствует действительности.
Сложный процесс оплодотворения происходит в верхней трети яйцепрово- да. Спермии быка непосредственно перед оплодотворением должны не менее 6 ч находиться в абдоминальной части яйцепровода. За это время они «созревают» и становятся способными оплодотворять. Такое явление называется капа- цитацией (инкубацией) спермы. У спермиев барана капацитация длится 1— 1,5 ч, а у хряка —2—3 ч. Оплодотворение может произойти только при достаточном скоплении спермиев. Количество их значительно колеблется даже у одного и того же вида животных. Спермии ожидают яйцеклетку. Последняя, как и спермии, должна «дозреть». Этот процесс также происходит в яйцепрово- де, куда яйцеклетка попадает в стадии овоцита. Там она при наличии активных спермиев выделяет направительные тельца и созревает. Сколько времени требуется для «дозревания» яйцеклетки, пока неясно. У овец, видимо, не более 2 ч.
Установлено, что для нормального процесса оплодотворения, а стало быть, получения жизнеспособного плода необходимо соединение только полноценных половых клеток. Неполноценные гаметы не вступают в процесс оплодотворения или вступают, но зародыши гибнут на ранней стадии. В связи с этим важное значение имеют время осеменения, возраст половых клеток и активность спермиев.
Оплодотворение у животных состоит из нескольких стадий.
| ОПЛОДОТВОРЕНИЕ И РАЗВИТИЕ ЗИГОТЫ |
Первая стадия — денудация — характеризуется тем, что яйцеклетка, проходя по яйцепроводу, освобождается от окружающих ее фолликулярных клеток (лучистого венца). Основная роль в денудации принадлежит спермиям, которые, внедряясь между клетками фолликулярного эпителия, выделяют ферменты, главным образом гиалуронидазу. За счет этих ферментов и неизвестных еще факторов разжижается
гиалуроновая кислота, входящая в состав студенистого вещества, связывающего клетки лучистого венца. Рассеивание клеток лучистого венца — это не видовая особенность, и оно может происходить под влиянием спермиев животных другого вида. Так, при осеменении свиней смесью спермы быка и хряка спермии быка освобождают яйцо от фолликулярных клеток, а проникают в него только спермии хряка. Процессу денудации способствуют также механические препятствия, обусловленные ворсинками слизистой оболочки яйце- провода.
Для оплодотворения не обязательно полное освобождение яйцеклетки от клеток лучистого венца. Достаточно лишь места, чтобы спермии проникли через прозрачную оболочку яйцеклетки в околожелточное пространство. Этот процесс, составляющий вторую стадию оплодотворения, в видовом отношении более специфичен. В это время заканчивается созревание яйцеклетки (выделение второго направительного тельца). Ядро, содержащее гаплоидное число хромосом, превращается в женский пронуклеус.
На третьей стадии оплодотворения один, реже несколько спермиев проникают через желточную оболочку яйцеклетки в ее цитоплазму. Это уже строго специфический процесс, поскольку в яйцеклетку могут проникать только спермии своего вида. При этом проникает не весь спермий, а лишь его головка и шейка. Внедрившись в цитоплазму яйцеклетки, спермий претерпевает большие изменения. Головка спермия быстро увеличивается в размере в десятки раз; вследствие ассимиляции цитоплазмы яйца она достигает величины ядра яйцеклетки и превращается в мужской пронуклеус, имеющий, как и женский пронуклеус, половинный набор хромосом.
На четвертой стадии про- нуклеусы (ядра яйцеклетки и спермия) постепенно сближаются, вступают в тесный контакт, быстро уменьшаются в объеме и полностью сливаются. Этот процесс, основной в оплодотворении, является еще недостаточно изученным. Образуется качественно новая клетка (зигота), ядро которой содержит диплоидное число хромосом. Образовавшаяся зигота, таким образом, имеет двойную наследственность. Она получает могучий стимул для дальнейшего развития и начинает быстро дробиться (рис. 40). После первого деления зиготы образуются два бластомера (дочерние клетки); оба бластомера дают четыре внучатых, а четыре внучатых — восемь правнучатых бластомеров и т. д. У млекопитающих такой закономерности нет: количество бластомеров у них даже в начальных стадиях не всегда бывает четным. В отличие от деления одноклеточных организмов клетки, образующиеся в результате деления зиготы, не расходятся, а остаются соединенными; в первый период общая величина зародыша не изменяется, поэтому деление зиготы называется дроблением. Дробление зиготы коровы длится около 8 сут. В течение первых 4 сут дробление происходит в яйцепро- воде, а затем в одном из рогов матки. Уже через 48 ч после овуляции зигота состоит из двух бластомеров, через 72 ч — из трех, через 84 ч — из шести и через 96 ч она поступает в стадии мору- лы в рог матки. Зигота по размеру равна яйцевой клетке или незначительно превышает ее, так как с каждым дроблением при увеличении количества бластомеров величина каждого из них соответственно уменьшается.
Наружный слой бластомеров, прилегающий к прозрачной оболочке, называется трофобластом (питающим листком); бластомеры, заключенные в трофобласт, образуют э м б - риобласт (зародышевыйлисток).
Полиспермия (переоплодотворение). Проникновение в цитоплазму яйцевой клетки двух или нескольких спермиев называется полиспермией. Из сотен миллионов спермиев, попадающих в половые пути самки при коитусе, лишь тысячи достигают бахромки.
В норме после проникновения в яйцо одного спермия другие не пропускаются в силу так называемой зональной реакции. У стареющих яиц зональная реакция нарушается, что обусловливает невозможность проникновения спермия в яйцо, а иногда под влиянием каких-то условий происходит слияние ядра яйца с несколькими спермиями. Это происходит, вероятно, тогда, когда приблизившиеся к яйцу спермии одновременно проникают внутрь его.
Полиспермия нарушает течение эмбриогенеза, приводит зиготу к гибели или, по мнению некоторых авторов, обусловливает развитие уродливых плодов (паразитизм, дицефализм и др.).
При физиологической полиспермии у млекопитающих спермии, не сливающиеся с ядром, по- видимому, только активизируют зиготу к дроблению аналогично реакции организма на введение инородного белка.
Множественное оплодотворение, или суперфекундация ($ирегГесипс1аПо). Это оплодотворение нескольких яйцевых клеток в период одного полового цикла, но спермой разных производителей. Оно часто наблюдается у многоплодных животных (собаки, свиньи, кошки), реже — у кобыл и коров. Суперфекундация, развившаяся в результате осеменения производителями, принадлежащими к одной породе и одинаковой с самкой масти, часто остается незамеченной; она выявляется, когда несколько производителей, принимавших участие в осеменении самки, принадлежит к разным породам и даже видам. Так, из- за недосмотра в стадию возбуждения у породистых собак наряду с чистопородными могут родиться щенки с признаками двух или трех пород, нескольких мастей и т. д. Отмечены случаи суперфе- кундации у свиней, когда рождаются поросята различных мастей; у кобыл, когда одновременно рождаются нормально развитые жеребенок и муленок.
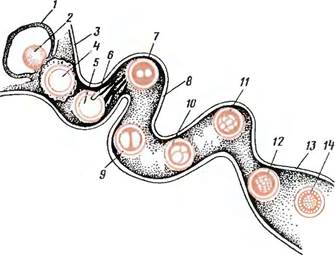 Рис. 40. Схема оплодотворения и продвижения зародыша по яйцепроводу в матку:
1— яичник; 2—лопнувший зрелый фолликул; 3—воронка яйцепровода; 4 —яйцос лучистым венцом; 5—яйцо, освобожденное от лучистого венца; 6 —проникновение спермия через прозрачную оболочку яйца; 7— образование зиготы (слияние мужского и женского пронуклеусов); 8—яйцепровод; 9— стадия двух бластомеров; 10—стадия четырех бла- стомеров; 11 —стадия восьми бластомеров; 12—стадия морулы; 13—верхушка рога матки; 14—стадия бластоцисты
Рис. 40. Схема оплодотворения и продвижения зародыша по яйцепроводу в матку:
1— яичник; 2—лопнувший зрелый фолликул; 3—воронка яйцепровода; 4 —яйцос лучистым венцом; 5—яйцо, освобожденное от лучистого венца; 6 —проникновение спермия через прозрачную оболочку яйца; 7— образование зиготы (слияние мужского и женского пронуклеусов); 8—яйцепровод; 9— стадия двух бластомеров; 10—стадия четырех бла- стомеров; 11 —стадия восьми бластомеров; 12—стадия морулы; 13—верхушка рога матки; 14—стадия бластоцисты
|
В настоящее время в животноводческой практике суперфекундацию применяют для повышения оплодотворяемое- ти и жизненности приплода. С этой целью при естественном осеменении практикуется коитус с двумя самцами с интервалом 10—15 мин.
Под трансплантацией зародышей понимают перенос зародышей из половых органов животных-доноров в половые органы животных-реципиентов. При этом в организме реципиентов наступает нормальная беременность с развитием зародышей, а затем и плодов.
Работы по трансплантации зародышей были начаты в конце XIX в. на кроликах с целью изучения развития плода и влияния организма матери на его развитие (Хипп, 1891). В. С. Груздев (1897) предпринял первые попытки по оплодотворению яйцеклеток и инкубации зародышей млекопитающих вне организма. В дальнейшем эти опыты продолжали многие отечественные и зарубежные ученые. А. В. Квасницкий впервые в мире (1949—1953 гг.) получил потомство у свиней после пересадок яйцеклеток и зигот. Под руководством А. И. Лопырина осуществлялись успешные пересадки зародышей овец. Появились публикации о пересадках оплодотворенных и неоплодотворенных яйцеклеток кроликов, коз, коров, лошадей. Практиковался хирургический метод получения и пересадки свежих зародышей.
С 1970 г. научные исследования и практическое применение трансплантации зародышей начали быстро развиваться. В 1971 г. проведены успешные опыты по замораживанию и оттаиванию зародышей мышей, а в 1973 г. родился первый теленок после пересадки корове замороженного и оттаянного зародыша.
За 10 лет (1973-1982 гг.) в Канаде получили 5413 телят-трансплантатов, а в США в 1990 г., когда они были в числе 35 стран (830 членов) Международного общества трансплантации зародышей (МОТЗ, 1984), - уже 500 тыс., в СССР к 1990 г. - 12 тыс.
В последние 12 лет в области трансплантации зародышей достигнуты рекордные результаты: в США от коровы- донора за год получено 136 телят, во
Франции — 80, в Германии — 57; в Российской Федерации в ВИЖ — от 44 доноров 261 теленок, на Украине в НИИЖЛиП — от 6 доноров 71 теленок.
В России образованы областные центры трансплантации зародышей и стабильно получают телят-трансплантатов в Воронеже (Ю. А. Скрипицын, 1992), Туле (1987), Омске (Т. А. Авдеева, 1994), а также создан республиканский центр трансплантации зародышей в Татарстане РЦТЗ (Р. М. Сайтов* 1995) и других республиках.
Современные методы суперовуляции позволяют увеличить в 10—20 раз число образующихся в яичниках яйцеклеток: у коров и овец — до 25, у свиней — до 40— 45, у лошадей —до 5, у грызунов —до 90. В Англии еще в 1983 г. от коровы за один раз извлекли 27 зародышей и, пересадив их реципиентам, получили 19 телят. Разрабатываются новые методы получения зародышей монозиготных близнецов (С. В. Вилладсон, Кэмб- ридж): двоен от коров, лошадей, свиней, овец, а в последние годы и из 4—8-кле- точных зародышей — четверен коров и овец. Получены 5 пар половинных однояйцовых близнецов и 46 близнецов из четвертых частей зародышей. За год от донора можно получать по 4—6 зачатий.
| ТРАНСПЛАНТАЦИЯ ЗАРОДЫШЕЙ |
Длительное сохранение зародышей позволяет создавать запасы (банки зародышей) ценного генетического фонда и экспортировать или импортировать зародышей высокопродуктивных и редких пород животных. Пересадка зародышей животных мясных пород молочным коровам имеет неоспоримое преимущество по сравнению со скрещиванием мясных быков с такими коровами и, следовательно, будет способствовать увеличению производства говядины. Возможно получение двоен путем пересадки по одному зародышу в каждый рог матки или введением дополнительного зародыша в небеременный рог матки. Можно получать ценный в пле-
Отбор

| 'и. оцененных по начестВи потом ства |
| синхронизация полаВпго цикла, реципиентов с полоВым шлам доноров |
тиНотныт-донораВ по Воспроизводительным качеством и продуктивности

|
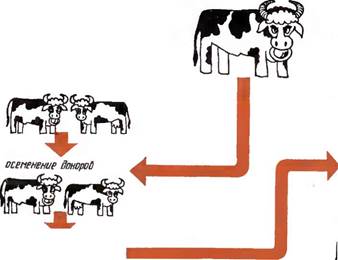
| гормональная оБраБотна с целью ВызВать множественно обуляцию |
| диагностика охоты |
| извлечение зародышей /их оценка нультиВираВа- ние, хранение/ |
| Рис. 41. Схема пересалки зародышей |

| лересодно зародышей решпиенлщ^ |
| Беременность и роды реципиентов, телят |
диагностика охоты
менном отношении приплод от коров местных пород, имеющих иммунитет к различным болезням.
Пересадку зародышей можно использовать и для изучения многих вопросов физиологии и патологии беременности, межвидовой гибридизации. Например, после пересадки овцам зародышей коз рождаются нормальные козлята, а от коз после трансплантации им зародышей овец — ягнята.
Однако преимущество от трансплантации зародышей в условиях хозяйства достигается только при четком повседневном осуществлении комплекса агро- зооветорганизационных мероприятий. Важнейший элемент его — наличие здоровых, конституционально крепких доноров и реципиентов, обеспеченных полноценным кормлением, хорошим содержанием и правильной эксплуатацией; высококвалифицированных акушеров-гинекологов; специального центра (пункта) трансплантации зародышей с необходимым оборудованием и оснащением.
Работу по трансплантации проводят в следующем порядке:
отбор доноров и реципиентов; вызывание множественной овуляции (суперовуляции) у доноров и их осеменение;
получение зародышей от доноров; оценка, культивирование и хранение зародышей;
синхронизация полового цикла реципиентов с половым циклом доноров;
пересадка зародышей на стадии мо- рулы или бластулы реципиентам (рис. 41).
Донор — это высокоценное, выдающееся животное, от которого после гормонального вызывания полиовуляции и осеменения спермой проверенного про- изводителя-улучшателя получают несколько зародышей. Отбирают только тех животных, которые обладают способностью к множественной овуляции и дают в течение длительного срока их использования большое количество зародышей, пригодных к пересадке. В качестве доноров лучше использовать здоровых коров в возрасте от 4 до 5 лет с хорошо развитой молочной железой, пригодной к машинному доению, и у которых не было каких-либо осложнений родов и послеродового периода. Первая стадия возбуждения полового цикла после родов должна быть синхронной и полноценной, с ярко выраженными феноменами: течки, полового возбуждения и охоты.
| ОТБОР ДОНОРОВ |
Операция пересадки зародышей экономически выгодна только в том случае, когда в качестве доноров берут выдающихся в племенном отношении животных. В некоторых случаях для получения зародышей рекомендуют использовать ценных в племенном отношении коров в заключительные сроки их продуктивной жизни, чтобы получить от них больше потомков.
ВЫЗЫВАНИЕ СУПЕРОВУЛЯЦИИ
Теоретические исследования и опыт проведения пересадок зародышей в производственных условиях показали, что для широкого распространения этого метода в практике необходимы надежные источники получения оплодотворенных яйцеклеток или зародышей на ранних стадиях развития.
В решении данной задачи имеются два принципиальных подхода. Первый подход — это обеспечение множественной овуляции, или полиовуляции, у коров, ремонтных телок и у молодых телочек до наступления половой зрелости; второй подход, находящийся пока в стадии разработки, — изыскание способов извлечения яйцеклеток непосредственно из яичников с дальнейшим созданием условий для их созревания и оплодотворения ш уИго. В настоящее время в практике используется в основном первый способ. Животным, обычно в середине полового цикла, вводят препараты гонадотропных гормонов, вызывающих усиленный рост и развитие многих пузырчатых фолликулов и их овуляцию на фоне обратного развития желтого тела.
Суперовуляцию считают достигнутой, если произошло выделение не менее трех яйцеклеток (в отдельных случаях у животных их овулирует 100 и более). Однако основная цель гормональной обработки — получение в результате суперовуляции 10—20 яйцеклеток.
У коров и телок для вызывания множественной овуляции применяют гона- дотропины гипофизарного и плацентарного происхождения; для обработки используют разнообразные схемы. Наиболее эффективны гонадотропины сыворотки жеребых кобыл (ГСЖК). Они вырабатываются в оболочках плода особыми клетками, которые внедряются в эндометрий кобылы примерно с 36-го по 40-й день беременности. Гонадотропины выделяются в кровь матери. Наиболее высокое содержание ГСЖК в крови кобыл отмечается примерно с 60-го по 90-й день беременности. Гонадотропины СЖК лучше всего применять в середине полового цикла: с 8-го по 15— 16-й день. Препараты вводят однократно в дозе от 2 тыс. до 3 тыс. ИЕ. Через 48 ч после введения ГСЖК инъецируют простагландин Ф2 (ПГФ2) или один из его синтетических аналогов. Обычно че
рез 2 дня наступает стадия возбуждения полового цикла с проявлением течки, общего возбуждения, охоты и овуляции. В этот период осеменяют животных-до- норов.
Преимуществом ГСЖК является их доступность (можно получать в необходимом количестве при наличии достаточного числа беременных кобыл). Но они медленно инактивируются в организме, период полураспада гонадотро- пинов в организме коров составляет 6 дней, и их еще можно обнаружить в крови через 10 дней после введения. Гона- дотропины после овуляции оказывают вредное влияние на качество развивающихся оплодотворенных яйцеклеток. При этом увеличивается возможность формирования фолликулярных кист яичников. Поэтому были предприняты попытки ограничить действие ГСЖК путем использования сыворотки, содержащей антитела. Антисыворотку к ГСЖК вводят донорам после суперову- ляторной обработки в начале проявления стадии возбуждения полового цикла, но результаты не всегда бывают надежными. Многие авторы считают, что благодаря введению антисыворотки у подопытных животных-доноров увеличивается количество овулирующих фолликулов по сравнению с контрольными животными, существенно уменьшается число фолликулов с кистозной дистрофией, а также наблюдается положительное влияние на оплодотворяемость овулирующих яйцеклеток.
С целью вызывания множественной овуляции можно применять препараты фолликулостимулирующего гормона (ФСГ). Они дают такую же эффективность, как ГСЖК, но их сложнее получать. Суперовуляцию можно вызывать ФСГ, получаемыми из гипофизов свиней и овец. Эти препараты имеют короткий период активности. Так, период полураспада ФСГ из гипофизов овец составляет 5 ч. По этой причине их назначают в снижающейся дозировке по 2 раза в день в течение 5 дней (первый и второй дни утром и вечером по 5 мг, третий день — соответственно по 4 мг, четвертый—по 3 мг и пятый — по 2 мг), всего Юдоз. Вводить ФСГ обычно начинают с 9-го по 11 -й день полового цикла. В ряде стран ФСГ используют в комбинации с лютеинизирующим гормоном (ЛГ) в соотношении 5:1 или в сочетании с простагландинами, которые инъецируют на третий день после начала обработки; в среднем через 48 ч после их введения животные-доноры проявляют стадию возбуждения полового цикла.
Ряд авторов сообщают, что после применения препаратов ФСГ получают больше зародышей, пригодных к пересадке, чем при использовании ГСЖК.
Некоторые авторы предлагают при использовании ФСГ упростить стандартную схему обработки и вместо двукратных проводить одноразовые инъекции в течение 5 дней. Этим достигается снижение затрат труда, уменьшение стрессорных влияний обработок на донора и повышается количество овуляций с 6,4 до 8,1.
При различных режимах обработки число овулировавших фолликулов у разных животных очень сильно варьирует. Это зависит от индивидуальных особенностей животных и многих других факторов. Описаны различия в чувствительности к ГСЖК у животных отдельных пород и линий крупного рогатого скота. При этом молочные породы проявляют меньшую реакцию, чем мясные. У телок наблюдается более высокая реакция, чем у коров.
Коровы-доноры должны всегда получать полноценный рацион. Имеются четкие доказательства, что голодание коров-доноров в период после введения ГСЖК заметно уменьшает количество овуляций. Исследования с помощью лапароскопа показали, что множественная овуляция у коров и телок является длительным процессом и после общепринятой обработки (ГСЖК, ФСГ + + ПГФ2) продолжается 24 ч и более. Если в одном яичнике происходит более 9—10 овуляций, то он настолько увеличивается, что фимбрии яйцепровода неспособны его окружить и часть яйцеклеток может попасть в брюшную полость. Имеются предположения, что использование предварительного анализа индивидуального гормонального фона у конкретных животных позволит лучше понять причины нарушений при супер- овуляторной реакции и, модифицируя гормональные обработки с учетом физиологического состояния животных, можно уменьшить вариабельность в количестве овуляций и снизить число не- реагирующих животных.
ОСЕМЕНЕНИЕ ДОНОРОВ
Для их осеменения берут сперму выдающихся быков-производителей, проверенных по качеству потомства и признанных улучшателями продуктивности. Имеются доказательства, что при использовании некоторых быков достигается более высокая степень оплодотворяемое™ коров-доноров, поэтому следует оценивать быков и по данному показателю. Отбор быков и работу со спермой проводят с соблюдением вете- ринарно-санитарных правил и согласно действующей инструкции по искусственному осеменению коров и телок. После гормональной обработки доноров у них с помощью быков-пробников выявляют половую охоту не менее 2 раз в день. Примерно у 10—12% животных признаки стадии возбуждения полового цикла не проявляются. Осеменение животных, у которых обнаружена охота, проводят несколько раз с 12-часовыми интервалами до ее окончания, иногда его повторяют 3—4 раза. В каждой дозе спермы должно быть не менее 40— 50 млн живых подвижных спермиев. Чаще используют способ осеменения с ректальной фиксацией шейки матки, сперму вводят в ее канал.
Некоторые зарубежные авторы предлагают вводить сперму в полость тела матки. Имеются также рекомендации вводить одну порцию спермы в левый, а вторую — в правый рог матки. Свежие спермии сохраняют жизнеспособность в половых путях самок дольше, чем замороженные и оттаянные. Поэтому при использовании свежей спермы в течение охоты можно проводить 1—2 осеменения. При этом достигается более высокая степень оплодотворяемости. Нецелесообразно осеменять доноров в отдаленные сроки после окончания охоты, поскольку в дальнейшем это может оказывать вредное влияние на степень извлечения пригодных к пересадке зародышей.
ИЗВЛЕЧЕНИЕ ЗАРОДЫШЕЙ
Оплодотворение яйцеклеток происходит в яйцепроводе. Образовавшиеся зиготы подвергаются дроблению, и большинство из них у крупного рогатого скота попадают в матку на 4-й день. Зародыши целесообразно извлекать у коров на 7—8-й день, а у овец — на 5—6-й день после первого осеменения (до освобождения зародыша из прозрачной оболочки). Для извлечения зародышей используют два способа: нехирургический и хирургический.
При нехирургическом способе извлечения зародышей животных фиксируют в станке. Прямую кишку освобождают от содержимого и проводят тщательное ректальное исследование. Определяют, сколько желтых тел находится в каждом яичнике. Хвост с помощью тесемки фиксируют. Проводят туалет и дезинфекцию наружных половых органов и промежности. Для прекращения перистальтики прямой кишки эпидурально вводят 10 мл 2%-ного раствора новокаина. Для вымывания зародышей из матки приме-
/ г
 Рис. 42. Катетер для извлечения зародышей:
1 — корпус катетера; 2 — мандрен; 3 — труба для нагнетания воздуха; 4—баллончик для воздуха
Рис. 42. Катетер для извлечения зародышей:
1 — корпус катетера; 2 — мандрен; 3 — труба для нагнетания воздуха; 4—баллончик для воздуха
|
няют различные инструменты. Большинство авторов используют гибкий од- ноходовой катетер Фолея с упругим мандреном и надувным баллончиком (рис. 42). Инструмент должен быть стерильным. Сперва катетер вводят во влагалище по верхнему его своду и проводят под ректальным контролем через канал шейки матки в рог матки (рис. 43). Для более полного извлечения зародышей нужно, не травмируя слизистую оболочку, как можно глубже ввести инструмент в рог матки. После того как катетер достигнет в роге матки необходимого положения, мандрен удаляют и в баллончик катетера накачивают 10— 15 мл воздуха. При этом катетер фиксируется в роге матки и промывная жидкость не вытекает мимо катетера.
Закрепив катетер, промывают полость рога матки с помощью шприца
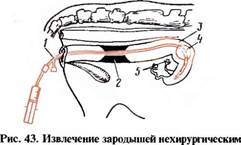 способом:
1 — катетер для извлечения зародышей; 2 — шейка и 3— рог матки; 4—надувной баллончик; 5 — яичники
способом:
1 — катетер для извлечения зародышей; 2 — шейка и 3— рог матки; 4—надувной баллончик; 5 — яичники
|
Люэра вместимостью 50—60 мл. В рог матки в зависимости от его величины вводят порциями от 40 до 60 мл промывной жидкости, затрачивая на промывание каждого рога не более 500 мл. Наполнение матки промывной средой и степень ее оттока контролируют рек- тально.
Для более полного извлечения зародышей верхушку рога матки приподнимают и выпрямляют. Некоторые авторы рекомендуют яйцепровод вблизи верхушки рога матки осторожно зажать большим и указательным пальцами. При этом предотвращается поступление в брюшную полость жидкости, содержащей зародыши. Однако практика показывает, что поступление в брюшную полость жидкости из рога матки отмечается только при наличии большого давления в матке, поэтому яйцепровод можно не зажимать. Перед извлечением катетера следует удалить воздух из баллончика. Таким же образом промывают и второй рог.
В качестве среды для промывания используют фосфатно-буферный солевой раствор (ФБС) Дюльбекко (табл. 13).
Раствор готовят на тридистиллиро- ванной воде. Первые 4 вещества растворяют в 800 мл, а 5-е и 6-е — каждое отдельно в 100 мл. В итоге получают три раствора, которые автоклавируют, а затем смешивают. В таком виде их можно хранить при 4 °С до 2 нед. Непосредственно перед употреблением в ФБС вводят следующие компоненты (в расчете на 1 л):
альбумин бычьей сыворотки — 4 г; глюкоза — 1 г (5,56 ммоль); Ка-пируват — 0,036 г (0,33 ммоль); пенициллин (калиевая соль) — 100 тыс. ЕД.
Собранную в цилиндр промывную жидкость отстаивают 20—35 мин при температуре 20—37 °С, чтобы зародыши опустились на дно, после чего верхний слой удаляют с помощью сифона. Нижний слой жидкости порционно по 20— 30 мл для обнаружения зародышей ис-

