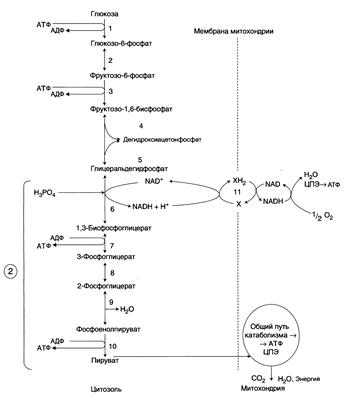
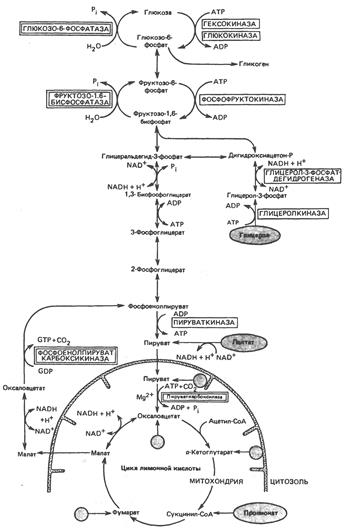
В аэробном процессе расщепления глюкозы выделяют 2 этапа:
1. Подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на 2 молекулы фосфотриоз Эта серия реакций протекает с использованием 2 молекул АТФ.
2. Этап, сопряжённый с синтезом АТФ. В результате этой серии реакций фосфотриозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10 моль АТФ.
Реакции аэробного расщепления глюкозы аналогичны таковым для гликолиза (анаэробного расщепления глюкозы).
Общая схема аэробного расщепления глюкозы:
1-10 – реакции аэробного расщепления глюкозы; 11 – малат-аспартатный челночный механизм транспорта водорода в митохондрии; 2 (в кружке) – стехиометрический коэффициент.
Общая схема гликолиза
Реакциями субстратного фосфорилирования являются реакции, в ходе которых происходит фосфорилирование АДФ (образование АТФ) за счёт отщепления остатков фосфорной кислоты от: 1,3-бисфосфоглицерата (фермент фосфоглицераткиназа) и фосфоенолпирувата (фермент пируваткиназа).
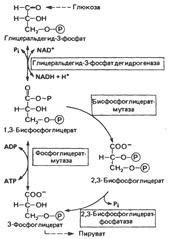
Реакции 2,3-бисфосфоглицератного цикла в эритроцитах
Малат-аспартатная челночная система.
1, 2 – окислительно-восстановительные реакции, обеспечивающие транспорт водорода из цитозоля в митохондрии на ЦПЭ; 3, 4 – транслоказы, обеспечивающие транспорт α-кетоглутарата, аспартата и глутамата через мембрану митохондрий.
Написать названия всех ферментов, катализирующих реакции аэробного расщепления глюкозы:
Реакции цикла Кребса и цепи дыхательных ферментов подробно описаны в лекции, посвящённой биоэнергетике.
4. Ключевые реакции глюконеогенеза. Аллостерическая регуляция ферментов гликолиза и глюконеогенеза.
Роль фруктозо-2,6-бисфосфата
Глюконеогенез – процесс синтеза глюкозы из веществ неуглеводной природы.
Основная функция глюконеогенеза – поддержание постоянного уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок.
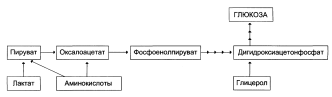
Глюконеогенез включает все механизмы и пути, обеспечивающие образование глюкозы и гликогена из неуглеводных компонентов. Главными субстратами глюконеогенеза служат глюкогенные аминокислоты, лактат, глицерол и (у жвачных) пропионат. Гликонеогенез происходит главным образом в печени и почках, поскольку именно в этих органах имеется полный набор необходимых ферментов.
Глюконеогенез обеспечивает потребности организма в глюкозе в тех случаях, когда диета содержит недостаточное количество углеводов. Постоянное поступление глюкозы в качестве источника энергии особенно необходимо для нервной системы и эритроцитов. При понижении концентрации глюкозы в крови ниже определенного критического уровня нарушается функционирование мозга; при тяжелой гипогликемии возникает коматозное состояние и может наступить летальный исход. Глюкоза необходима также для жировой ткани как источник глицерола, входящего в состав глицеридов; она играет, вероятно, существенную роль в поддержании эффективных концентраций интермедиатов цикла лимонной кислоты во многих тканях. Из этого следует, что даже в условиях, когда большая часть потребностей организма в калориях обеспечивается за счет жира, всегда сохраняется определенная потребность в глюкозе. Кроме того, глюкоза служит единственным видом топлива для работы скелетной мышцы в анаэробных условиях. Она является предшественником молочного сахара (лактозы) в молочных железах и активно потребляется плодом в период развития. Следует отметить также, что механизм глюконеогенеза используется для удаления из крови продуктов тканевого метаболизма, например лактата, образующегося в мышцах и эритроцитах, глицерола, непрерывно образующегося в жировой ткани.
Пропионат – главная глюкогенная жирная кислота, образующаяся в процессе переваривания углеводов жвачными животными, является главным субстратом глюконеогенеза у этих животных.
Метаболические пути, участвующие в глюконеогенезе, являются модификациями путей гликолиза и цикла лимонной кислоты. Кребс отметил, что простому обращению гликолиза препятствуют энергетические барьеры на ряде стадий:
1) между пируватом и фосфоенолпируватом,
2) между фруктозо-1,6-бисфосфатом и фруктозо-6-фосфатом,
3) между глюкозо-6-фосфатом и глюкозой,
4) между глюкозо-1 -фосфатом и гликогеном.
Эти барьеры обходятся с помощью специальных реакций.
1) В митохондриях имеется фермент пируваткарбоксилаза, который при участии АТР, биотина (витамина группы В) и СО2 превращает пируват в оксалоацетат. Функция биотина заключается в присоединении СО2 (из бикарбоната) к ферменту, далее СО2 переносится на пируват. Во внемитохондриальной среде клетки имеется второй фермент – фосфоенолпнруваткарбоксикиназа, который катализирует превращение оксалоацетата в фосфоенолпируват. Для этой реакции требуется высокоэнергетический фосфат в форме GTP или ITP; в результате реакции освобождается СО2. Таким образом, с помощью этих двух ферментов и лактатдегидрогеназы лактат может превращаться в фосфоенолпируват.
Существенное препятствие, однако, заключается в том, что выход оксалоацетата из митохондрии весьма затруднен. Оно преодолевается следующим образом: оксалоацетат превращается в соединение, легко диффундирующее из митохондрии во внемитохондриальный компартмент клетки, где это соединение снова превращается в оксалоацетат. Таким соединением служит малат; его образование из оксалоацетата внутри митохондрий и превращение обратно в оксалоацетат вне митохондрий катализируются малатдегидрогеназой.
2) Превращение фруктозо-1,6-бисфосфата во фруктозо-6-фосфат, необходимое для обращения гликолиза на рассматриваемой стадии, катализируется специфическим ферментом фруктозо-1,6-бисфосфатазой. Это – ключевой фермент в том смысле, что именно его присутствием определяется, способна ли ткань ресинтезировать гликоген из пирувата и триозофосфатов. Этот фермент имеется в печени и почках, он был также обнаружен в поперечнополосатых мышцах. Считают, что в сердечной мышце и гладких мышцах он отсутствует.
3) Превращение глюкозо-6-фосфата в глюкозу катализируется другой специфической фосфатазой – глюкозо-6-фосфатазой. Она присутствует в печени и почках, но отсутствует в мышцах и жировой ткани. Наличие этого фермента позволяет ткани поставлять глюкозу в кровь.
4) Распад гликогена с образованием глюкозо-1-фосфата осуществляется фосфорилазой. Синтез гликогена идет по совершенно другому пути, через образование уридиндифосфатглюкозы, и катализируется гликогенсинтазой.
Переключение печени с гликолиза на глюконеогенез и обратно происходит с участием инсулина и глюкагона и осуществляется с помощью:
- аллостерической регуляции активности ферментов;
- ковалентной модификации ферментов путём фосфорилирования/дефосфорилирования;
- индукции/репрессии синтеза ключевых ферментов.
«Субстратные» циклы – парные комбинации процессов синтеза и распада метаболитов.
Направление реакций первого субстратного цикла регулируется главным образом концентрацией глюкозы.
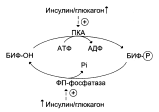
Направление реакций второго субстратного цикла зависит от активности фосфофруктокиназы и фосфатазы фруктозо-1,6-бисфосфата. Активность этих ферментов зависит от концентрации фруктозо-2,6-бисфосфата. Фруктозо-2,6-бисфосфат – метабоилит, образующийся в незначительных количествах из фруктозо-6-фосфата и выполняющий только регуляторные функции. Образование фруктозо-2,6-бисфосфата путём фосфорилирования фруктозо-6-фосфата катализирует бифункциональный фермент (БИФ), который катализирует и обратную реакцию:
Киназная активность БИФ проявляется, когда фермент находится в дефосфорилированной форме (БИФ-ОН). Дефосфорилированная форма БИФ характерна для абсорбтивного периода, когда инсулин/глюкагоновый индекс высокий. В этот период количество фруктозо-2,6-бисфосфата увеличивается:
При низком инсулин/глюкагоновом индексе, характерном для периода длительного голодания, происходит фосфорилирование БИФ, и он функционирует как фосфатаза. Результат – снижение количества фруктозо-2,6-бисфосфата.
Регуляторное значение фруктозо-2,6-бисфосфата заключается в том, что он аллостерически активирует фосфофруктокиназу (фермент гликолиза). При этом фруктозо-2,6-бисфосфат снижает ингибирующее действие АТФ на этот фермент в абсорбтивном периоде и повышает его сродство к фруктозо-6-фосфату. В то же время фруктозо-2,6-бисфосфат ингибирует фруктозо-1,6-бисфосфатазу (фермент глюконеогенеза). Таким образом, в абсорбтивном периоде благодаря фруктозо-2,6-бисфосфату ускоряется процесс гликолиза.
Результатом уменьшения количества фруктозо-2,6-бисфосфата в постабсорбтивном периоде будет снижение активности фосфофруктокиназы, замедление гликолиза и переключение гликолиза на глюконеогенез.
Включение субстратов в глюконеогенез
Главные пути глюконеогенсза в печени
Кружок со стрелкой показывает места вступления в цикл глюкогенных аминокислот после переаминирования. Названия ключевых ферментов глюконсогенеза заключены в двойной прямоугольник. АТР, необходимый для поддержания глюконеогенеза, генерируется в ходе окисления ацетил-СоА, который в свою очередь образуется из длинноцепочечных жирных кислот или из лактата (через пируват. при действии пируватдегидрогеназы). У жвачных животных важным источником ацстил-СоА служит пропионат.
Регуляция реакций второго субстратного цикла фруктозо-2,6-бисфосфатом:
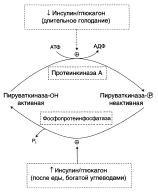
Р регуляции третьего субстратного цикла основная роль принадлежит пируваткиназе, фосфорилированная форма которой неактивна, а дефосфорилированная – активна:
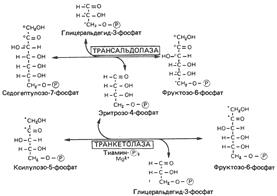
5. Реакции пентозофосфатного пути превращения глюкозы. Образование восстановительных эквивалентов и рибозы.
Пентозофосфатный путь
Пентозофосфатный цикл не приводит к синтезу АТР, он выполняет две главные функции:
1) образование NADPH для восстановительных синтезов, таких, как синтез жирных кислот и стероидов;
2) обеспечение рибозой синтеза нуклеотидов и нуклеиновых кислот. Недостаточность ряда ферментов пентозофосфатного пути является причиной гемолиза эритроцитов. Например, одна из форм гемолитической анемии обусловлена недостаточностью глюкозо-6-фосфатдегидрогеназы.
Метаболическое значение пентозофосфатного пути
Сравнение с гликолизом
Пентозофосфатный путь существенно отличается от гликолиза. Окисление осуществляется на первой стадии, и в нем участвует не NAD, как в гликолизе, a NADP; одним.из продуктов пентозофосфатного пути является СО2, который в реакциях гликолиза не образуется. Наконец, пентозофосфатный путь не генерирует АТР.
Образование восстановительных эквивалентов
Значение метаболического пути для различных тканей можно оценить по его активности. Пентозофосфатный путь активно протекает в печени, жировой ткани, коре надпочечников, щитовидной железе, эритроцитах, семенниках и в молочных железах в период лактации; он неактивен в нелактирующей молочной железе и малоактивен в скелетных мышцах. Все ткани, в которых активность данного пути высока, используют в реакциях восстановительного синтеза NADPH, например в реакциях синтеза жирных кислот, стероидов, аминокислот (с участием глутаматдегидрогеназы) или восстановленного глутатиона в эритроцитах. Вероятно, в условиях активного липогенеза или при наличии любой системы, утилизирующей NADPH, возрастает активная деградация глюкозы по пентозофосфатному пути в связи с увеличением отношения NADP: NADPH. В условиях, которые возникают после приема пищи, может индуцироваться синтез глюкозо-6-фосфат-дегидрогеназы и 6-фосфоглюконатдегидрогеназы.
Образование рибозы
Пентозофосфатный путь поставляет рибозу для синтеза нуклеотидов и нуклеиновых кислот. Источником рибозы является интермедиат рибозо-5-фосфат, который в реакции с АТР образует PRPP, используемый в биосинтезе нуклеотидов. Мышечная ткань содержит очень малые количества глюкозо-6-фосфатдегидрогеназы и 6-фосфоглюконатдегидрогеназы. Тем не менее скелетная мышца способна синтезировать рибозу. Вероятно, это осуществляется при обращении неокислительной фазы пентозофосфатного пути, утилизирующей фруктозо-6-фосфат. Таким образом, синтез рибозы может осуществляться в ткани, если в ней протекает часть реакций пентозофосфатного пути.
6. Метаболизм фруктозы и галактозы. Регуляция уровня глюкозы в крови. Источники глюкозы крови. Цикл Кори и глюкозо-аланиновый цикл. Почечный порог для глюкозы, глюкозурия. Толерантность к глюкозе
Источники глюкозы крови:
1. Глюкоза пищи.
2. Гликоген.
3. Лактат (цикл Кори).
4. Аминокислоты (глюкозо-аланиновый цикл).
5. Глицерол.
Путь превращения галактозы в глюкозу и путь синтеза лактозы
Клинические аспекты
Нарушение метаболизма галактозы наблюдается при галактоземии, которая может быть вызвана наследственными дефектами в любом из трех ферментов. Наиболее хорошо известным является недостаток уридилтрансферазы. При увеличении концентрации галактозы в крови повышается ее концентрация в тканях. В тканях глаза она восстанавливается альдозоредуктазой с образованием соответствующего полиола (галактитола). Накопление галактитола способствует развитию катаракты. Весьма тяжелые последствия наблюдаются при дефиците уридилтрансферазы: в печени происходит накопление галактозо-1-фосфата, при этом соответственно снижается концентрация неорганического фосфата. В результате возникает нарушение функции печени, а затем расстройство психики. Если при наследственном дефиците галактозо-1-фосфат-уридилтрансферазы, приводящем к нарушению метаболизма галактозы в печени и красных кровяных тельцах, эпимераза присутствует в достаточном количестве, то у больных может происходить образование UDP-галактозы из глюкозы. Это объясняет, почему дети с таким заболеванием могут нормально расти и развиваться при назначении диеты, из которой исключена галактоза (такая диета назначается для предотвращения тяжелых форм заболевания). Описано несколько различных генетических дефектов, которые вызывают не полный, а частичный дефицит трансферазы. Поскольку обычно этот фермент присутствует в организме в избытке, снижение его активности до 50% (и даже ниже) может не сопровождаться клиническими проявлениями заболевания; последние наблюдаются у гомозиготных индивидуумов. В тех случаях, когда имеется дефицит эпимеразы в эритроцитах, при наличии данного фермента в печени и других органах симптомы заболевания не обнаруживаются.
Регуляция концентрации глюкозы в крови
Поддержание содержания глюкозы в крови на определенном уровне является примером одного из самых совершенных механизмов гомеостаза, в функционировании которого участвуют печень, внепеченочные ткани и некоторые гормоны. Глюкоза легко проникает в клетки печени и относительно медленно в клетки внепеченочных тканей. Следовательно, прохождение через клеточную мембрану является лимитирующей скорость стадией при потреблении глюкозы внепеченочными тканями. Поступившая в клетки глюкоза быстро фосфорилируется при действии гексокиназы. С другой стороны, вполне возможно, что более значительное влияние на потребление глюкозы печенью или на выход глюкозы из этого органа оказывают активность некоторых других ферментов и концентрации ключевых промежуточных продуктов. Тем не менее концентрация глюкозы в крови является важным фактором, регулирующим скорость потребления глюкозы как печенью, так и внепеченочными тканями.
Роль глюкокнназы
Следует особо отметить, что глюкозо-6-фосфат ингибирует гексокиназу и, следовательно, потребление глюкозы внепеченочными тканями, зависящее от гексокиназы, которая катализирует фосфорилирование глюкозы и регулируется по принципу обратной связи. С печенью этого не происходит, поскольку глюкозо-6-фосфат не ингибирует глюкокиназу. Этот фермент характеризуется более высоким значением Км (более низким сродством) для глюкозы, чем гексокиназа; активность глюкокиназы повышается в пределах физиологических концентраций глюкозы; после приема богатой углеводами пищи фермент «настраивается» на высокие концентрации глюкозы, поступающей в печень по воротной вене. Отметим, что этот фермент отсутствует у жвачных, у которых лишь небольшое количество глюкозы поступает из кишечника в систему воротной вены.
При нормальном содержании глюкозы в крови (80-100 мг/100 мл) печень, по-видимому, поставляет глюкозу в кровь. При увеличении же уровня глюкозы в крови ее выход из печени прекращается, а при достаточно высоких концентрациях начинается поступление глюкозы в печень. Как показали опыты, проведенные на крысах, при концентрации глюкозы в воротной вене печени 150 мг/100 мл скорость поступления глюкозы в печень и скорость ее выхода из печени равны.
Роль инсулина
В состоянии гипергликемии увеличивается поступление глюкозы как в печень, так и в периферические ткани. Центральную роль в регуляции концентрации глюкозы в крови играет гормон - инсулин. Он синтезируется в поджелудочной железе В-клетками островков Лангерганса, и его поступление в кровь увеличивается при гипергликемии. Концентрация этого гормона в крови изменяется параллельно концентрации глюкозы; введение его быстро вызывает гипогликемию. К веществам, вызывающим секрецию инсулина, относятся аминокислоты, свободные жирные кислоты, кетоновые тела, глюкагон, секретин и лекарственный препарат толбутамид; адреналин и норадреналин, наоборот, блокируют его секрецию. Инсулин быстро вызывает увеличение поглощения глюкозы жировой тканью и мышцами за счет ускорения транспорта глюкозы через клеточные мембраны путем перемещения переносчиков глюкозы из цитоплазмы в плазматическую мембрану. Однако инсулин не оказывает прямого действия на проникновение глюкозы в клетки печени; это согласуется с данными о том, что скорость метаболизма глюкозы клетками печени не лимитируется скоростью ее прохождения через клеточные мембраны. Инсулин, однако, действует опосредованно, влияя на активность ферментов, участвующих в гликолизе и гликогенолизе (см. выше).
Передняя доля гипофиза секретирует гормоны, действие которых противоположно действию инсулина, т. е. они повышают уровень глюкозы в крови. К ним относятся гормон роста, АКТГ (кортикотропин) и, вероятно, другие «диабетогенные» факторы. Гипогликемия стимулирует секрецию гормона роста. Он вызывает уменьшение поступления глюкозы в некоторые ткани, например в мышцы. Действие гормона роста является до некоторой степени опосредованным, поскольку он стимулирует мобилизацию из жировой ткани свободных жирных кислот, которые являются ингибиторами потребления глюкозы. Длительное введение гормона роста приводит к диабету. Вызывая гипергликемию, он стимулирует постоянную секрецию инсулина, что в конечном счете приводит к истощению В-клеток.
Глюкокортнконды (11-гидроксистероиды) секретируются корой надпочечников и играют важную роль в углеводном обмене. Введение этих стероидов усиливает глюконеогенез за счет интенсификации катаболизма белков в тканях, увеличения потребления аминокислот печенью, а также повышения активности трансаминаз и других ферментов, участвующих в процессе глюконеогенеза в печени. Кроме того, глюкокортикоиды ингибируют утилизацию глюкозы во внепеченочных тканях. В рассмотренных случаях глюкокортикоиды действуют подобно антагонистам инсулнна.
Адреналин секретируется мозговым слоем надпочечников в ответ на стрессорные стимулы (страх, сильное волнение, кровотечение, кислородная недостаточность, гипогликемия и т. д.). Стимулируя фосфорилазу, он вызывает гликогенолиз в печени и мышцах. В мышцах из-за отсутствия глюкозо-6-фосфатазы гликогенолиз доходит до стадии лактата, в то время как в печени основным продуктом превращения гликогена является глюкоза, которая поступает в кровь, где уровень ее повышается.
Глюкагон является гормоном, секретируемым А-клетками островков Лангерганса в поджелудочной железе (его секреция стимулируется гипогликемией). Когда по воротной вене глюкагон поступает в печень, он, подобно адреналину, активирует фосфорилазу и вызывает гликогенолиз. Большая часть эндогенного глюкагона задерживается в печени. В отличие от адреналина глюкагон не влияет на фосфорилазу мышц. Этот гормон усиливает также глюконеогенез из аминокислот и лактата. Гипергликемический эффект глюкагона обусловлен как гликогенолизом, так и глюконеогенезом в печени.
Следует отметить, что гормон щитовидной железы также влияет на содержание глюкозы в крови. Экспериментальные данные свидетельствуют о том, что тироксин обладает диабетогенным действием, а удаление щитовидной железы препятствует развитию диабета. Было отмечено, что гликоген полностью отсутствует в печени животных с тиреотоксикозом. У людей с усиленной функцией щитовидной железы содержание сахара в крови при голодании повышено, а у людей с пониженной функцией щитовидной железы оно снижено. При гипертиреозе глюкоза, по-видимому, расходуется с нормальной или повышенной скоростью, а при гипотиреозе способность утилизировать глюкозу понижена. Следует отметить, что пациенты с гипофункцией щитовидной железы менее чувствительны к действию инсулина, чем здоровые люди и пациенты с гипертиреозом.
Глюкозо-аланиновый цикл
Цикл Кори (глюкозо-лактатный цикл)
1 – поступление лактата из сокращающейся мышцы с током крови в печень;
2 – синтез глюкозы из лактата в печени;
3 – поступление глюкозы из печени с током крови в работающую мышцу;
4 – использование глюкозы как энергетического субстрата сокращающейся мышцей и образование лактата.
Концентрация глюкозы в крови
У человека в период между приемами пищи концентрация глюкозы в крови варьирует от 80 до 100 мг/100 мл. После приема пищи, богатой углеводами, концентрация глюкозы увеличивается до 120-130 мг/100 мл. Во время голодания концентрация глюкозы падает приблизительно до 60-70 мг/100 мл. При нормальном состоянии организма уровень глюкозы в крови колеблется в указанных пределах.
Метаболизм фруктозы
Альдолаза А обнаружена во всех тканях, кроме печени, в которой имеется только
альдолаза В. Альдозоредуктаза не обнаружена в печени.
Почечный порог для глюкозы, глюкозурия
Когда содержание глюкозы в крови достигает относительно высокого уровня, в процесс регуляции включаются и почки. Глюкоза фильтруется почечными клубочками и обычно полностью возвращается в кровь в результате реабсорбции (обратного всасывания) в почечных канальцах. Процесс реабсорбции глюкозы связан с расходованием АТР в клетках почечных канальцев. Максимальная скорость реабсорбции глюкозы в почечных канальцах около 350 мг·мин-1. При повышенном содержании глюкозы в крови клубочковый фильтрат содержит больше глюкозы, чем может быть реабсорбировано в канальцах. Избыток глюкозы выводится с мочой, т.е. возникает гликозурия. У здоровых людей гликозурия наблюдается в том случае, если содержание глюкозы в венозной крови превышает 170-180 мг/100 мл; этот уровень называют почечным порогом для глюкозы.
У подопытных животных можно вызвать гликозурию с помощью флоридзина, ингибирующего реабсорбцию глюкозы в почечных канальцах. Такую гликозурию, обусловленную нарушением реабсорбции глюкозы, называют почечной гликозурией. Причиной почечной гликозурии может быть наследственный дефект почек, или же она может развиться в результате ряда заболеваний. Гликозурия часто является указанием на заболевание сахарным диабетом.
Толерантность к глюкозе
О способности организма использовать глюкозу можно судить по его толерантности к ней. После введения определенного количества глюкозы строят кривые динамики содержания глюкозы в крови, которые характеризуют толерантность к глюкозе. При сахарном диабете она понижена из-за уменьшения количества секретируемого инсулина; при этом заболевании содержание глюкозы в крови повышается (гипергликемия), возникает гликозурия, могут происходить изменения в обмене жиров. Толерантность к глюкозе снижается не только при диабете, но и при некоторых состояниях, сопровождающихся нарушением функции печени, при ряде инфекционных заболеваний, ожирении, действии ряда лекарственных препаратов, а иногда и при атеросклерозе. Снижение толерантности к глюкозе может также наблюдаться при гиперфункции гипофиза или коры надпочечников вследствие антагонизма между гормонами, секретируемыми этими железами внутренней секреции, и инсулином.
Инсулин повышает толерантность организма к глюкозе. При его введении содержание глюкозы в крови снижается, а ее потребление и содержание в виде гликогена в печени и мышцах увеличиваются. При введении избытка инсулина может возникнуть тяжелая гипогликемия, сопровождающаяся судорогами; если в этом состоянии быстро не ввести глюкозу, то может наступить летальный исход. У человека гипогликемические судороги появляются при быстром снижении содержания глюкозы в крови до 20 мг/100 мл. Повышенная толерантность к глюкозе наблюдается при недостаточной функции гипофиза или коры надпочечников; это является следствием снижения антагонистического эффекта гормонов, секретируемых этими железами, по отношению к инсулину. В результате «относительное содержание» инсулина в организме увеличивается.
Проба на толерантность к глюкозе
Кривые содержания глюкозы в крови у здорового и больного диабетом человека после приема 50 г глюкозы. Обратите внимание на то, что у больного диабетом исходное содержание глюкозы в крови повышено. Показателем нормальной толерантности является возвращение к исходному уровню глюкозы в крови в течение двух часов.





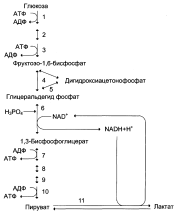 Реакциями субстратного фосфорилирования являются реакции, в ходе которых происходит фосфорилирование АДФ (образование АТФ) за счёт отщепления остатков фосфорной кислоты от: 1,3-бисфосфоглицерата (фермент фосфоглицераткиназа) и фосфоенолпирувата (фермент пируваткиназа).
Реакции 2,3-бисфосфоглицератного цикла в эритроцитах
Реакциями субстратного фосфорилирования являются реакции, в ходе которых происходит фосфорилирование АДФ (образование АТФ) за счёт отщепления остатков фосфорной кислоты от: 1,3-бисфосфоглицерата (фермент фосфоглицераткиназа) и фосфоенолпирувата (фермент пируваткиназа).
Реакции 2,3-бисфосфоглицератного цикла в эритроцитах
 Малат-аспартатная челночная система.
Малат-аспартатная челночная система.
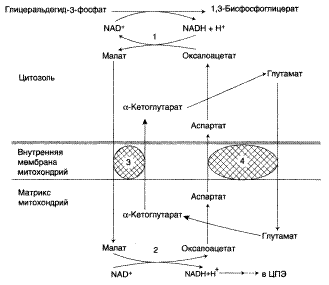 1, 2 – окислительно-восстановительные реакции, обеспечивающие транспорт водорода из цитозоля в митохондрии на ЦПЭ; 3, 4 – транслоказы, обеспечивающие транспорт α-кетоглутарата, аспартата и глутамата через мембрану митохондрий.
Написать названия всех ферментов, катализирующих реакции аэробного расщепления глюкозы:
Реакции цикла Кребса и цепи дыхательных ферментов подробно описаны в лекции, посвящённой биоэнергетике.
1, 2 – окислительно-восстановительные реакции, обеспечивающие транспорт водорода из цитозоля в митохондрии на ЦПЭ; 3, 4 – транслоказы, обеспечивающие транспорт α-кетоглутарата, аспартата и глутамата через мембрану митохондрий.
Написать названия всех ферментов, катализирующих реакции аэробного расщепления глюкозы:
Реакции цикла Кребса и цепи дыхательных ферментов подробно описаны в лекции, посвящённой биоэнергетике.
 Киназная активность БИФ проявляется, когда фермент находится в дефосфорилированной форме (БИФ-ОН). Дефосфорилированная форма БИФ характерна для абсорбтивного периода, когда инсулин/глюкагоновый индекс высокий. В этот период количество фруктозо-2,6-бисфосфата увеличивается:
Киназная активность БИФ проявляется, когда фермент находится в дефосфорилированной форме (БИФ-ОН). Дефосфорилированная форма БИФ характерна для абсорбтивного периода, когда инсулин/глюкагоновый индекс высокий. В этот период количество фруктозо-2,6-бисфосфата увеличивается:
 При низком инсулин/глюкагоновом индексе, характерном для периода длительного голодания, происходит фосфорилирование БИФ, и он функционирует как фосфатаза. Результат – снижение количества фруктозо-2,6-бисфосфата.
Регуляторное значение фруктозо-2,6-бисфосфата заключается в том, что он аллостерически активирует фосфофруктокиназу (фермент гликолиза). При этом фруктозо-2,6-бисфосфат снижает ингибирующее действие АТФ на этот фермент в абсорбтивном периоде и повышает его сродство к фруктозо-6-фосфату. В то же время фруктозо-2,6-бисфосфат ингибирует фруктозо-1,6-бисфосфатазу (фермент глюконеогенеза). Таким образом, в абсорбтивном периоде благодаря фруктозо-2,6-бисфосфату ускоряется процесс гликолиза.
Результатом уменьшения количества фруктозо-2,6-бисфосфата в постабсорбтивном периоде будет снижение активности фосфофруктокиназы, замедление гликолиза и переключение гликолиза на глюконеогенез.
При низком инсулин/глюкагоновом индексе, характерном для периода длительного голодания, происходит фосфорилирование БИФ, и он функционирует как фосфатаза. Результат – снижение количества фруктозо-2,6-бисфосфата.
Регуляторное значение фруктозо-2,6-бисфосфата заключается в том, что он аллостерически активирует фосфофруктокиназу (фермент гликолиза). При этом фруктозо-2,6-бисфосфат снижает ингибирующее действие АТФ на этот фермент в абсорбтивном периоде и повышает его сродство к фруктозо-6-фосфату. В то же время фруктозо-2,6-бисфосфат ингибирует фруктозо-1,6-бисфосфатазу (фермент глюконеогенеза). Таким образом, в абсорбтивном периоде благодаря фруктозо-2,6-бисфосфату ускоряется процесс гликолиза.
Результатом уменьшения количества фруктозо-2,6-бисфосфата в постабсорбтивном периоде будет снижение активности фосфофруктокиназы, замедление гликолиза и переключение гликолиза на глюконеогенез.
 Главные пути глюконеогенсза в печени
Главные пути глюконеогенсза в печени
 Кружок со стрелкой показывает места вступления в цикл глюкогенных аминокислот после переаминирования. Названия ключевых ферментов глюконсогенеза заключены в двойной прямоугольник. АТР, необходимый для поддержания глюконеогенеза, генерируется в ходе окисления ацетил-СоА, который в свою очередь образуется из длинноцепочечных жирных кислот или из лактата (через пируват. при действии пируватдегидрогеназы). У жвачных животных важным источником ацстил-СоА служит пропионат.
Регуляция реакций второго субстратного цикла фруктозо-2,6-бисфосфатом:
Кружок со стрелкой показывает места вступления в цикл глюкогенных аминокислот после переаминирования. Названия ключевых ферментов глюконсогенеза заключены в двойной прямоугольник. АТР, необходимый для поддержания глюконеогенеза, генерируется в ходе окисления ацетил-СоА, который в свою очередь образуется из длинноцепочечных жирных кислот или из лактата (через пируват. при действии пируватдегидрогеназы). У жвачных животных важным источником ацстил-СоА служит пропионат.
Регуляция реакций второго субстратного цикла фруктозо-2,6-бисфосфатом:
 Р регуляции третьего субстратного цикла основная роль принадлежит пируваткиназе, фосфорилированная форма которой неактивна, а дефосфорилированная – активна:
Р регуляции третьего субстратного цикла основная роль принадлежит пируваткиназе, фосфорилированная форма которой неактивна, а дефосфорилированная – активна:
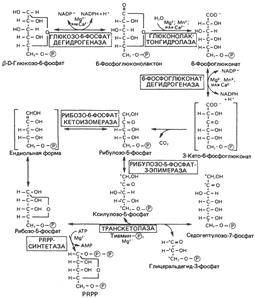
 PRPP – 5-фосфорибозил-1-пирофосфат.
PRPP – 5-фосфорибозил-1-пирофосфат.
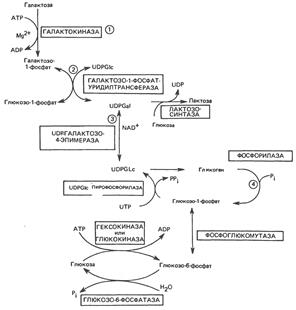 Клинические аспекты
Нарушение метаболизма галактозы наблюдается при галактоземии, которая может быть вызвана наследственными дефектами в любом из трех ферментов. Наиболее хорошо известным является недостаток уридилтрансферазы. При увеличении концентрации галактозы в крови повышается ее концентрация в тканях. В тканях глаза она восстанавливается альдозоредуктазой с образованием соответствующего полиола (галактитола). Накопление галактитола способствует развитию катаракты. Весьма тяжелые последствия наблюдаются при дефиците уридилтрансферазы: в печени происходит накопление галактозо-1-фосфата, при этом соответственно снижается концентрация неорганического фосфата. В результате возникает нарушение функции печени, а затем расстройство психики. Если при наследственном дефиците галактозо-1-фосфат-уридилтрансферазы, приводящем к нарушению метаболизма галактозы в печени и красных кровяных тельцах, эпимераза присутствует в достаточном количестве, то у больных может происходить образование UDP-галактозы из глюкозы. Это объясняет, почему дети с таким заболеванием могут нормально расти и развиваться при назначении диеты, из которой исключена галактоза (такая диета назначается для предотвращения тяжелых форм заболевания). Описано несколько различных генетических дефектов, которые вызывают не полный, а частичный дефицит трансферазы. Поскольку обычно этот фермент присутствует в организме в избытке, снижение его активности до 50% (и даже ниже) может не сопровождаться клиническими проявлениями заболевания; последние наблюдаются у гомозиготных индивидуумов. В тех случаях, когда имеется дефицит эпимеразы в эритроцитах, при наличии данного фермента в печени и других органах симптомы заболевания не обнаруживаются.
Регуляция концентрации глюкозы в крови
Поддержание содержания глюкозы в крови на определенном уровне является примером одного из самых совершенных механизмов гомеостаза, в функционировании которого участвуют печень, внепеченочные ткани и некоторые гормоны. Глюкоза легко проникает в клетки печени и относительно медленно в клетки внепеченочных тканей. Следовательно, прохождение через клеточную мембрану является лимитирующей скорость стадией при потреблении глюкозы внепеченочными тканями. Поступившая в клетки глюкоза быстро фосфорилируется при действии гексокиназы. С другой стороны, вполне возможно, что более значительное влияние на потребление глюкозы печенью или на выход глюкозы из этого органа оказывают активность некоторых других ферментов и концентрации ключевых промежуточных продуктов. Тем не менее концентрация глюкозы в крови является важным фактором, регулирующим скорость потребления глюкозы как печенью, так и внепеченочными тканями.
Роль глюкокнназы
Следует особо отметить, что глюкозо-6-фосфат ингибирует гексокиназу и, следовательно, потребление глюкозы внепеченочными тканями, зависящее от гексокиназы, которая катализирует фосфорилирование глюкозы и регулируется по принципу обратной связи. С печенью этого не происходит, поскольку глюкозо-6-фосфат не ингибирует глюкокиназу. Этот фермент характеризуется более высоким значением Км (более низким сродством) для глюкозы, чем гексокиназа; активность глюкокиназы повышается в пределах физиологических концентраций глюкозы; после приема богатой углеводами пищи фермент «настраивается» на высокие концентрации глюкозы, поступающей в печень по воротной вене. Отметим, что этот фермент отсутствует у жвачных, у которых лишь небольшое количество глюкозы поступает из кишечника в систему воротной вены.
При нормальном содержании глюкозы в крови (80-100 мг/100 мл) печень, по-видимому, поставляет глюкозу в кровь. При увеличении же уровня глюкозы в крови ее выход из печени прекращается, а при достаточно высоких концентрациях начинается поступление глюкозы в печень. Как показали опыты, проведенные на крысах, при концентрации глюкозы в воротной вене печени 150 мг/100 мл скорость поступления глюкозы в печень и скорость ее выхода из печени равны.
Роль инсулина
В состоянии гипергликемии увеличивается поступление глюкозы как в печень, так и в периферические ткани. Центральную роль в регуляции концентрации глюкозы в крови играет гормон - инсулин. Он синтезируется в поджелудочной железе В-клетками островков Лангерганса, и его поступление в кровь увеличивается при гипергликемии. Концентрация этого гормона в крови изменяется параллельно концентрации глюкозы; введение его быстро вызывает гипогликемию. К веществам, вызывающим секрецию инсулина, относятся аминокислоты, свободные жирные кислоты, кетоновые тела, глюкагон, секретин и лекарственный препарат толбутамид; адреналин и норадреналин, наоборот, блокируют его секрецию. Инсулин быстро вызывает увеличение поглощения глюкозы жировой тканью и мышцами за счет ускорения транспорта глюкозы через клеточные мембраны путем перемещения переносчиков глюкозы из цитоплазмы в плазматическую мембрану. Однако инсулин не оказывает прямого действия на проникновение глюкозы в клетки печени; это согласуется с данными о том, что скорость метаболизма глюкозы клетками печени не лимитируется скоростью ее прохождения через клеточные мембраны. Инсулин, однако, действует опосредованно, влияя на активность ферментов, участвующих в гликолизе и гликогенолизе (см. выше).
Передняя доля гипофиза секретирует гормоны, действие которых противоположно действию инсулина, т. е. они повышают уровень глюкозы в крови. К ним относятся гормон роста, АКТГ (кортикотропин) и, вероятно, другие «диабетогенные» факторы. Гипогликемия стимулирует секрецию гормона роста. Он вызывает уменьшение поступления глюкозы в некоторые ткани, например в мышцы. Действие гормона роста является до некоторой степени опосредованным, поскольку он стимулирует мобилизацию из жировой ткани свободных жирных кислот, которые являются ингибиторами потребления глюкозы. Длительное введение гормона роста приводит к диабету. Вызывая гипергликемию, он стимулирует постоянную секрецию инсулина, что в конечном счете приводит к истощению В-клеток.
Глюкокортнконды (11-гидроксистероиды) секретируются корой надпочечников и играют важную роль в углеводном обмене. Введение этих стероидов усиливает глюконеогенез за счет интенсификации катаболизма белков в тканях, увеличения потребления аминокислот печенью, а также повышения активности трансаминаз и других ферментов, участвующих в процессе глюконеогенеза в печени. Кроме того, глюкокортикоиды ингибируют утилизацию глюкозы во внепеченочных тканях. В рассмотренных случаях глюкокортикоиды действуют подобно антагонистам инсулнна.
Адреналин секретируется мозговым слоем надпочечников в ответ на стрессорные стимулы (страх, сильное волнение, кровотечение, кислородная недостаточность, гипогликемия и т. д.). Стимулируя фосфорилазу, он вызывает гликогенолиз в печени и мышцах. В мышцах из-за отсутствия глюкозо-6-фосфатазы гликогенолиз доходит до стадии лактата, в то время как в печени основным продуктом превращения гликогена является глюкоза, которая поступает в кровь, где уровень ее повышается.
Глюкагон является гормоном, секретируемым А-клетками островков Лангерганса в поджелудочной железе (его секреция стимулируется гипогликемией). Когда по воротной вене глюкагон поступает в печень, он, подобно адреналину, активирует фосфорилазу и вызывает гликогенолиз. Большая часть эндогенного глюкагона задерживается в печени. В отличие от адреналина глюкагон не влияет на фосфорилазу мышц. Этот гормон усиливает также глюконеогенез из аминокислот и лактата. Гипергликемический эффект глюкагона обусловлен как гликогенолизом, так и глюконеогенезом в печени.
Следует отметить, что гормон щитовидной железы также влияет на содержание глюкозы в крови. Экспериментальные данные свидетельствуют о том, что тироксин обладает диабетогенным действием, а удаление щитовидной железы препятствует развитию диабета. Было отмечено, что гликоген полностью отсутствует в печени животных с тиреотоксикозом. У людей с усиленной функцией щитовидной железы содержание сахара в крови при голодании повышено, а у людей с пониженной функцией щитовидной железы оно снижено. При гипертиреозе глюкоза, по-видимому, расходуется с нормальной или повышенной скоростью, а при гипотиреозе способность утилизировать глюкозу понижена. Следует отметить, что пациенты с гипофункцией щитовидной железы менее чувствительны к действию инсулина, чем здоровые люди и пациенты с гипертиреозом.
Клинические аспекты
Нарушение метаболизма галактозы наблюдается при галактоземии, которая может быть вызвана наследственными дефектами в любом из трех ферментов. Наиболее хорошо известным является недостаток уридилтрансферазы. При увеличении концентрации галактозы в крови повышается ее концентрация в тканях. В тканях глаза она восстанавливается альдозоредуктазой с образованием соответствующего полиола (галактитола). Накопление галактитола способствует развитию катаракты. Весьма тяжелые последствия наблюдаются при дефиците уридилтрансферазы: в печени происходит накопление галактозо-1-фосфата, при этом соответственно снижается концентрация неорганического фосфата. В результате возникает нарушение функции печени, а затем расстройство психики. Если при наследственном дефиците галактозо-1-фосфат-уридилтрансферазы, приводящем к нарушению метаболизма галактозы в печени и красных кровяных тельцах, эпимераза присутствует в достаточном количестве, то у больных может происходить образование UDP-галактозы из глюкозы. Это объясняет, почему дети с таким заболеванием могут нормально расти и развиваться при назначении диеты, из которой исключена галактоза (такая диета назначается для предотвращения тяжелых форм заболевания). Описано несколько различных генетических дефектов, которые вызывают не полный, а частичный дефицит трансферазы. Поскольку обычно этот фермент присутствует в организме в избытке, снижение его активности до 50% (и даже ниже) может не сопровождаться клиническими проявлениями заболевания; последние наблюдаются у гомозиготных индивидуумов. В тех случаях, когда имеется дефицит эпимеразы в эритроцитах, при наличии данного фермента в печени и других органах симптомы заболевания не обнаруживаются.
Регуляция концентрации глюкозы в крови
Поддержание содержания глюкозы в крови на определенном уровне является примером одного из самых совершенных механизмов гомеостаза, в функционировании которого участвуют печень, внепеченочные ткани и некоторые гормоны. Глюкоза легко проникает в клетки печени и относительно медленно в клетки внепеченочных тканей. Следовательно, прохождение через клеточную мембрану является лимитирующей скорость стадией при потреблении глюкозы внепеченочными тканями. Поступившая в клетки глюкоза быстро фосфорилируется при действии гексокиназы. С другой стороны, вполне возможно, что более значительное влияние на потребление глюкозы печенью или на выход глюкозы из этого органа оказывают активность некоторых других ферментов и концентрации ключевых промежуточных продуктов. Тем не менее концентрация глюкозы в крови является важным фактором, регулирующим скорость потребления глюкозы как печенью, так и внепеченочными тканями.
Роль глюкокнназы
Следует особо отметить, что глюкозо-6-фосфат ингибирует гексокиназу и, следовательно, потребление глюкозы внепеченочными тканями, зависящее от гексокиназы, которая катализирует фосфорилирование глюкозы и регулируется по принципу обратной связи. С печенью этого не происходит, поскольку глюкозо-6-фосфат не ингибирует глюкокиназу. Этот фермент характеризуется более высоким значением Км (более низким сродством) для глюкозы, чем гексокиназа; активность глюкокиназы повышается в пределах физиологических концентраций глюкозы; после приема богатой углеводами пищи фермент «настраивается» на высокие концентрации глюкозы, поступающей в печень по воротной вене. Отметим, что этот фермент отсутствует у жвачных, у которых лишь небольшое количество глюкозы поступает из кишечника в систему воротной вены.
При нормальном содержании глюкозы в крови (80-100 мг/100 мл) печень, по-видимому, поставляет глюкозу в кровь. При увеличении же уровня глюкозы в крови ее выход из печени прекращается, а при достаточно высоких концентрациях начинается поступление глюкозы в печень. Как показали опыты, проведенные на крысах, при концентрации глюкозы в воротной вене печени 150 мг/100 мл скорость поступления глюкозы в печень и скорость ее выхода из печени равны.
Роль инсулина
В состоянии гипергликемии увеличивается поступление глюкозы как в печень, так и в периферические ткани. Центральную роль в регуляции концентрации глюкозы в крови играет гормон - инсулин. Он синтезируется в поджелудочной железе В-клетками островков Лангерганса, и его поступление в кровь увеличивается при гипергликемии. Концентрация этого гормона в крови изменяется параллельно концентрации глюкозы; введение его быстро вызывает гипогликемию. К веществам, вызывающим секрецию инсулина, относятся аминокислоты, свободные жирные кислоты, кетоновые тела, глюкагон, секретин и лекарственный препарат толбутамид; адреналин и норадреналин, наоборот, блокируют его секрецию. Инсулин быстро вызывает увеличение поглощения глюкозы жировой тканью и мышцами за счет ускорения транспорта глюкозы через клеточные мембраны путем перемещения переносчиков глюкозы из цитоплазмы в плазматическую мембрану. Однако инсулин не оказывает прямого действия на проникновение глюкозы в клетки печени; это согласуется с данными о том, что скорость метаболизма глюкозы клетками печени не лимитируется скоростью ее прохождения через клеточные мембраны. Инсулин, однако, действует опосредованно, влияя на активность ферментов, участвующих в гликолизе и гликогенолизе (см. выше).
Передняя доля гипофиза секретирует гормоны, действие которых противоположно действию инсулина, т. е. они повышают уровень глюкозы в крови. К ним относятся гормон роста, АКТГ (кортикотропин) и, вероятно, другие «диабетогенные» факторы. Гипогликемия стимулирует секрецию гормона роста. Он вызывает уменьшение поступления глюкозы в некоторые ткани, например в мышцы. Действие гормона роста является до некоторой степени опосредованным, поскольку он стимулирует мобилизацию из жировой ткани свободных жирных кислот, которые являются ингибиторами потребления глюкозы. Длительное введение гормона роста приводит к диабету. Вызывая гипергликемию, он стимулирует постоянную секрецию инсулина, что в конечном счете приводит к истощению В-клеток.
Глюкокортнконды (11-гидроксистероиды) секретируются корой надпочечников и играют важную роль в углеводном обмене. Введение этих стероидов усиливает глюконеогенез за счет интенсификации катаболизма белков в тканях, увеличения потребления аминокислот печенью, а также повышения активности трансаминаз и других ферментов, участвующих в процессе глюконеогенеза в печени. Кроме того, глюкокортикоиды ингибируют утилизацию глюкозы во внепеченочных тканях. В рассмотренных случаях глюкокортикоиды действуют подобно антагонистам инсулнна.
Адреналин секретируется мозговым слоем надпочечников в ответ на стрессорные стимулы (страх, сильное волнение, кровотечение, кислородная недостаточность, гипогликемия и т. д.). Стимулируя фосфорилазу, он вызывает гликогенолиз в печени и мышцах. В мышцах из-за отсутствия глюкозо-6-фосфатазы гликогенолиз доходит до стадии лактата, в то время как в печени основным продуктом превращения гликогена является глюкоза, которая поступает в кровь, где уровень ее повышается.
Глюкагон является гормоном, секретируемым А-клетками островков Лангерганса в поджелудочной железе (его секреция стимулируется гипогликемией). Когда по воротной вене глюкагон поступает в печень, он, подобно адреналину, активирует фосфорилазу и вызывает гликогенолиз. Большая часть эндогенного глюкагона задерживается в печени. В отличие от адреналина глюкагон не влияет на фосфорилазу мышц. Этот гормон усиливает также глюконеогенез из аминокислот и лактата. Гипергликемический эффект глюкагона обусловлен как гликогенолизом, так и глюконеогенезом в печени.
Следует отметить, что гормон щитовидной железы также влияет на содержание глюкозы в крови. Экспериментальные данные свидетельствуют о том, что тироксин обладает диабетогенным действием, а удаление щитовидной железы препятствует развитию диабета. Было отмечено, что гликоген полностью отсутствует в печени животных с тиреотоксикозом. У людей с усиленной функцией щитовидной железы содержание сахара в крови при голодании повышено, а у людей с пониженной функцией щитовидной железы оно снижено. При гипертиреозе глюкоза, по-видимому, расходуется с нормальной или повышенной скоростью, а при гипотиреозе способность утилизировать глюкозу понижена. Следует отметить, что пациенты с гипофункцией щитовидной железы менее чувствительны к действию инсулина, чем здоровые люди и пациенты с гипертиреозом.
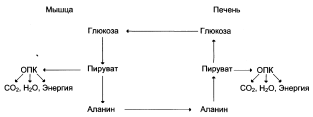 Цикл Кори (глюкозо-лактатный цикл)
Цикл Кори (глюкозо-лактатный цикл)
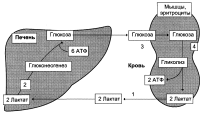 1 – поступление лактата из сокращающейся мышцы с током крови в печень;
2 – синтез глюкозы из лактата в печени;
3 – поступление глюкозы из печени с током крови в работающую мышцу;
4 – использование глюкозы как энергетического субстрата сокращающейся мышцей и образование лактата.
Концентрация глюкозы в крови
У человека в период между приемами пищи концентрация глюкозы в крови варьирует от 80 до 100 мг/100 мл. После приема пищи, богатой углеводами, концентрация глюкозы увеличивается до 120-130 мг/100 мл. Во время голодания концентрация глюкозы падает приблизительно до 60-70 мг/100 мл. При нормальном состоянии организма уровень глюкозы в крови колеблется в указанных пределах.
Метаболизм фруктозы
1 – поступление лактата из сокращающейся мышцы с током крови в печень;
2 – синтез глюкозы из лактата в печени;
3 – поступление глюкозы из печени с током крови в работающую мышцу;
4 – использование глюкозы как энергетического субстрата сокращающейся мышцей и образование лактата.
Концентрация глюкозы в крови
У человека в период между приемами пищи концентрация глюкозы в крови варьирует от 80 до 100 мг/100 мл. После приема пищи, богатой углеводами, концентрация глюкозы увеличивается до 120-130 мг/100 мл. Во время голодания концентрация глюкозы падает приблизительно до 60-70 мг/100 мл. При нормальном состоянии организма уровень глюкозы в крови колеблется в указанных пределах.
Метаболизм фруктозы
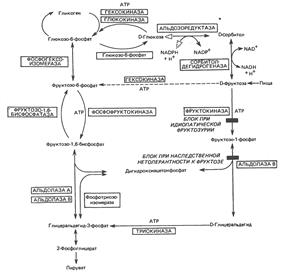 Альдолаза А обнаружена во всех тканях, кроме печени, в которой имеется только
альдолаза В. Альдозоредуктаза не обнаружена в печени.
Почечный порог для глюкозы, глюкозурия
Когда содержание глюкозы в крови достигает относительно высокого уровня, в процесс регуляции включаются и почки. Глюкоза фильтруется почечными клубочками и обычно полностью возвращается в кровь в результате реабсорбции (обратного всасывания) в почечных канальцах. Процесс реабсорбции глюкозы связан с расходованием АТР в клетках почечных канальцев. Максимальная скорость реабсорбции глюкозы в почечных канальцах около 350 мг·мин-1. При повышенном содержании глюкозы в крови клубочковый фильтрат содержит больше глюкозы, чем может быть реабсорбировано в канальцах. Избыток глюкозы выводится с мочой, т.е. возникает гликозурия. У здоровых людей гликозурия наблюдается в том случае, если содержание глюкозы в венозной крови превышает 170-180 мг/100 мл; этот уровень называют почечным порогом для глюкозы.
У подопытных животных можно вызвать гликозурию с помощью флоридзина, ингибирующего реабсорбцию глюкозы в почечных канальцах. Такую гликозурию, обусловленную нарушением реабсорбции глюкозы, называют почечной гликозурией. Причиной почечной гликозурии может быть наследственный дефект почек, или же она может развиться в результате ряда заболеваний. Гликозурия часто является указанием на заболевание сахарным диабетом.
Толерантность к глюкозе
О способности организма использовать глюкозу можно судить по его толерантности к ней. После введения определенного количества глюкозы строят кривые динамики содержания глюкозы в крови, которые характеризуют толерантность к глюкозе. При сахарном диабете она понижена из-за уменьшения количества секретируемого инсулина; при этом заболевании содержание глюкозы в крови повышается (гипергликемия), возникает гликозурия, могут происходить изменения в обмене жиров. Толерантность к глюкозе снижается не только при диабете, но и при некоторых состояниях, сопровождающихся нарушением функции печени, при ряде инфекционных заболеваний, ожирении, действии ряда лекарственных препаратов, а иногда и при атеросклерозе. Снижение толерантности к глюкозе может также наблюдаться при гиперфункции гипофиза или коры надпочечников вследствие антагонизма между гормонами, секретируемыми этими железами внутренней секреции, и инсулином.
Инсулин повышает толерантность организма к глюкозе. При его введении содержание глюкозы в крови снижается, а ее потребление и содержание в виде гликогена в печени и мышцах увеличиваются. При введении избытка инсулина может возникнуть тяжелая гипогликемия, сопровождающаяся судорогами; если в этом состоянии быстро не ввести глюкозу, то может наступить летальный исход. У человека гипогликемические судороги появляются при быстром снижении содержания глюкозы в крови до 20 мг/100 мл. Повышенная толерантность к глюкозе наблюдается при недостаточной функции гипофиза или коры надпочечников; это является следствием снижения антагонистического эффекта гормонов, секретируемых этими железами, по отношению к инсулину. В результате «относительное содержание» инсулина в организме увеличивается.
Проба на толерантность к глюкозе
Альдолаза А обнаружена во всех тканях, кроме печени, в которой имеется только
альдолаза В. Альдозоредуктаза не обнаружена в печени.
Почечный порог для глюкозы, глюкозурия
Когда содержание глюкозы в крови достигает относительно высокого уровня, в процесс регуляции включаются и почки. Глюкоза фильтруется почечными клубочками и обычно полностью возвращается в кровь в результате реабсорбции (обратного всасывания) в почечных канальцах. Процесс реабсорбции глюкозы связан с расходованием АТР в клетках почечных канальцев. Максимальная скорость реабсорбции глюкозы в почечных канальцах около 350 мг·мин-1. При повышенном содержании глюкозы в крови клубочковый фильтрат содержит больше глюкозы, чем может быть реабсорбировано в канальцах. Избыток глюкозы выводится с мочой, т.е. возникает гликозурия. У здоровых людей гликозурия наблюдается в том случае, если содержание глюкозы в венозной крови превышает 170-180 мг/100 мл; этот уровень называют почечным порогом для глюкозы.
У подопытных животных можно вызвать гликозурию с помощью флоридзина, ингибирующего реабсорбцию глюкозы в почечных канальцах. Такую гликозурию, обусловленную нарушением реабсорбции глюкозы, называют почечной гликозурией. Причиной почечной гликозурии может быть наследственный дефект почек, или же она может развиться в результате ряда заболеваний. Гликозурия часто является указанием на заболевание сахарным диабетом.
Толерантность к глюкозе
О способности организма использовать глюкозу можно судить по его толерантности к ней. После введения определенного количества глюкозы строят кривые динамики содержания глюкозы в крови, которые характеризуют толерантность к глюкозе. При сахарном диабете она понижена из-за уменьшения количества секретируемого инсулина; при этом заболевании содержание глюкозы в крови повышается (гипергликемия), возникает гликозурия, могут происходить изменения в обмене жиров. Толерантность к глюкозе снижается не только при диабете, но и при некоторых состояниях, сопровождающихся нарушением функции печени, при ряде инфекционных заболеваний, ожирении, действии ряда лекарственных препаратов, а иногда и при атеросклерозе. Снижение толерантности к глюкозе может также наблюдаться при гиперфункции гипофиза или коры надпочечников вследствие антагонизма между гормонами, секретируемыми этими железами внутренней секреции, и инсулином.
Инсулин повышает толерантность организма к глюкозе. При его введении содержание глюкозы в крови снижается, а ее потребление и содержание в виде гликогена в печени и мышцах увеличиваются. При введении избытка инсулина может возникнуть тяжелая гипогликемия, сопровождающаяся судорогами; если в этом состоянии быстро не ввести глюкозу, то может наступить летальный исход. У человека гипогликемические судороги появляются при быстром снижении содержания глюкозы в крови до 20 мг/100 мл. Повышенная толерантность к глюкозе наблюдается при недостаточной функции гипофиза или коры надпочечников; это является следствием снижения антагонистического эффекта гормонов, секретируемых этими железами, по отношению к инсулину. В результате «относительное содержание» инсулина в организме увеличивается.
Проба на толерантность к глюкозе
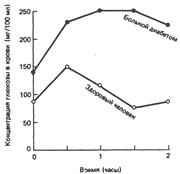 Кривые содержания глюкозы в крови у здорового и больного диабетом человека после приема 50 г глюкозы. Обратите внимание на то, что у больного диабетом исходное содержание глюкозы в крови повышено. Показателем нормальной толерантности является возвращение к исходному уровню глюкозы в крови в течение двух часов.
Кривые содержания глюкозы в крови у здорового и больного диабетом человека после приема 50 г глюкозы. Обратите внимание на то, что у больного диабетом исходное содержание глюкозы в крови повышено. Показателем нормальной толерантности является возвращение к исходному уровню глюкозы в крови в течение двух часов.

