




Структурные элементы мужской репродуктивной системы связаны между собой нейрогормональной регуляцией, что позволяет ей функционировать как единое целое. Нейрогормональные механизмы регуляции мужской репродуктивной системы сложны. Они включают ЦНС, которая через нейротрансмиттеры регулирует функцию гипоталамуса. Как известно, гипоталамус содержит пептидергические и моноаминергические нейросекреторные клетки. Эти элементы, составляющие фундаментальную основу механизмов регуляции функций половых желез, теснейшим образом взаимодействуют и, в свою очередь, находятся под регулирующим влиянием нейропроводников внегипоталамических образований головного мозга [81]. В аркаунтных ядрах медиабазального гипоталамуса синтезируется гонадотропин-рилизинг-гормон, который секретируется в портальную систему (гонадолиберин или люлиберин) и стимулирует продукцию двух гормонов гипофиза – ЛГ и ФСГ. Последние оказывают стимулирующее действие на семенники (яички). Расчеты на крысах показали, что каждая молекула гонадолюберина, достигнувшая аденогипофиза через систему сосудов гипофизарного стебля, индуцирует выделение 50–60 молекул ЛГ [82]. Семенники, в свою очередь, вырабатывают половые гормоны, которые оказывают тормозящее действие на секрецию гонадотропных гормонов (рис. 2.5). Таким образом, гормональные механизмы регуляции мужской репродуктивной системы обеспечивают состояние физиологического равновесия между гонадотропинами плазмы и половыми стероидами [83].
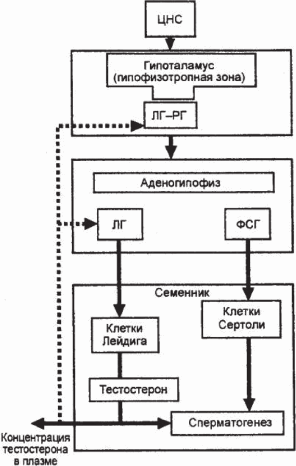
Рис. 2.5. Регуляция уровня тестостерона и процесса сперматогенеза: рилизинг фактор лютеинизирующего гормона (ЛГ – РГ) – гонадолиберин; сплошными стрелками обозначено усиление секреции, пунктирными – торможение по типу обратной связи
Основные половые стероиды мужского организма (андрогены) синтезируются в яичках (семенниках). Вещества с андрогенной активностью в небольшом количестве продуцирует также кора надпочечников. Среди них можно отметить следующие: андростерон, 4-андростен-3,17-дион, дегидроизоандростерон и др. Каждая ткань, продуцирующая стероиды, имеет свой собственный характерный профиль продуктов секреции. Все стероиды липофильны, это означает их низкую способность растворяться в воде. Поэтому в крови 95 % стероидных гормонов находятся в связанном состоянии со специфическими транспортными белками. Равновесие между связанными и свободными стероидами подчиняется закону действующих масс. С помощью транспортных белков гормоны переносятся к своим органам-мишеням. Только свободные, не связанные с белком стероиды являются биологически активными. Секс-стероидсвязывающий глобулин (СССГ) специфично связывает эстрадиол и андрогены с низкой емкостью и высокой аффинностью, в то время как кортико-стероидсвязываюший глобулин (КСГ) связывает прогестерон и глюко-кортикоиды. Помимо своей транспортной функции специфические сывороточные гормон-связывающие белки защищают стероиды от метаболической инактивации по пути от секретирующей их железы к органу-мишени. СССГ и КСГ являются кислыми гликопротеидами с молекулярной массой 45 тыс. и 65 тыс. соответственно.
2.3.1. Мужские половые гормоны – андрогены. Биосинтез и метаболизм гормонов яичек (семенников)
Широкое развитие исследование андрогенов получило в 30-х годах ХХ в. В 1931 г. А. Бутенандту (Германия) удалось получить 15 мг андростенона из 10 тыс. л мочи. Этот анаболический стероид был также синтезирован Л. Ружичкой (Швейцария) в 1934 г. К этому времени уже было известно, что в экстракте яичек содержится более активный мужской половой стероидный гормон, чем андростенон. В 1935 г. Э. Лакер из Амстердама, переработав огромное количество семенников быков, получил 10 г чистого кристаллического тестостерона. Гормон получил такое название, исходя из своего происхождения: тесто – яичко, стерол – стероидная структура, и окончание – он, которое говорит о том, что вещество является кетоном. Химический синтез тестостерона из холестерола был произведен независимо друг от друга в этом же году А. Бутенандтом и Л. Ружичкой. За работы по половым гормонам эти ученые были удостоены в 1939 г. Нобелевской премии в области химии [84, 85].
Биологическая активность тестостерона оказалась в 10 раз выше таковой известного к тому времени андростерона. На основании ряда исследований было высказано предположение, что тестостерон является 17-дегидро-производным андростендиона. Вскоре гипотетическая структура тестостерона была расшифрована, а его синтез явился прологом для синтеза десятков производных тестостерона с заданными свойствами.
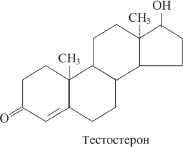
Мужские половые стероидные гормоны – андрогены (от греч. аndrоs – мужчина, genesis – происхождение) синтезируются в гормон-производящих клетках Лейдига, расположенных в интерстициальной ткани семенников между извитыми семенными канальцами. Они образуются также в надпочечниках и в небольшом количестве в яичниках. В тканях, продуцирующих стероиды, кроме тестостерона образуется еще пять важнейших андрогенов: дигидроэпиандростерон, адреностерон, Δ4-андростендион-3,17, 11β-окси-Δ4-андростендион-3,17, 11β-оксиэпиандростерон [86–89].
Основной мужской половой стероидный гормон – тестостерон. Он обусловливает как нормальный рост органов мужской репродуктивной системы, так и развитие вторичных половых признаков у мужчин. Он является антагонистом эстрадиола и резко снижает выделение азота с мочой, т. е. основной метаболический эффект тестостерона и других андрогенов – анаболический, который сопровождается усилением процессов синтеза нуклеиновых кислот, белка в клетках, некоторых ферментов и благодаря этому влияет практически на все виды обменных процессов в организме. Тестостерон, проникая в половые клетки, стимулирует развитие сперматоцитов в сперматиды, которые затем превращаются в сперматозоиды [81, 90].
У мужчин ежедневно вырабатывается 4-14 мг тестостерона, а его концентрация в плазме крови составляет 4,5–8,5 нг/мл (16–35 нмоль/л). Тестостерон преимущественно синтезируется в семенниках в интерстициальных клетках Лейдига (до 95 % от общего количества), которые располагаются между семенными канальцами. В корковом слое надпочечников, где также продуцируются андрогены, образование тестостерона крайне низкое, здесь синтезируются низкоактивные андрогены – дигидроэпиандростерон и андростендион. Их активность составляет не более 2–4 и 10 % соответственно от активности тестостерона. Тестостерон и другие андрогены образуются также у женщин в жировой ткани, печени, в коре надпочечников и в яичниках.
Циркулирующий тестостерон примерно на 98 % связан с белками плазмы, преимущественно с β-глобулином, связывающим половые стероиды. Этот белок, называемый обычно секс-гормон-связывающим глобулином (СГСГ) или тестостерон-эстрадиол-связывающим глобулином (ТЭСГ), образуется в печени. Основная функция СГСГ состоит, вероятно, в том, чтобы ограничивать концентрацию свободного тестостерона в сыворотке и перенос гормонов к своим органам-мишеням. Только свободные, не связанные с белком стероиды обладают биологическим действием. Всего в циркулирующей крови содержится около 2 % свободного тестостерона. Свободный тестостерон способен проникать в клетку, связываться с внутриклеточными рецепторами, проникать в ядро, изменять генную транскрипцию и в конечном счете реализовывать свои биологические эффекты. Помимо своей транспортной функции, гормонсвязывающие белки защищают стероиды от метаболической инактивации по пути от секретирующей железы к органу-мишени [87, 88, 91, 92].
Тестостерон поддерживает сперматогенез, стимулирует рост и функционирование добавочных половых желез, внешних половых органов. Гормон обладает анаболическим эффектом, главным образом, в отношении костей и мышц, с его участием происходят активация и синтез эритропоэтина в почках, стимуляция эритропоэза. У самцов это главный андроген, обусловливающий достижение половой зрелости. Достаточный уровень тестостерона представляет собой необходимое условие для осуществления половой функции у самцов [74, 91, 92].
Концентрация тестостерона в сыворотке крови в течение дня претерпевает изменения. У взрослых мужчин пик содержания тестостерона приходится на утреннее время и снижается к вечеру минимум на 25 %. После 30–40 лет отмечается прогрессирующее снижение уровня тестостерона в сыворотке крови на 1–2% в год. Это состояние получило название возрастного дефицита андрогенов.
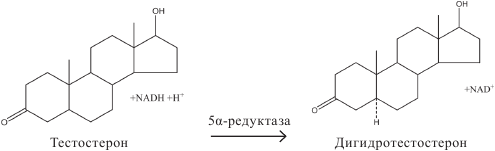
Тестостерон подвергается различным превращениям, среди которых самым важным является образование биологически активного 5α-дигидротестостерона (ДГТ) под влиянием фермента 5α-редуктазы в тканях-мишенях, т. е. вне семенников. В плазме взрослых мужчин содержание ДГТ примерно в 10 раз меньше содержания тестостерона: за сутки его образуется около 400 мкг. Из тестостерона образуется также другой высокоактивный андроген – андростандион. Небольшое количество тестостерона ароматизируется с образованием эстрадиола.
Главные 17-кетостероидные метаболиты тестостерона – андростерон и этиохоланолон – конъюгируют в печени с глюкоронидом и сульфатом с образованием водорастворимых соединений, которые не обладают биологической активностью и выводятся из организма.

