




Поскольку при данном виде транспорта энергия транспортируемого вещества уменьшается, то его перенос идёт только в одном направлении по концентрационному или электрохимическому градиенту. В соответствии с законом Фика диффузия прямо пропорциональная градиенту концентрации вещества, площади диффузионной мембраны, коэффициенту распределения и обратно пропорциональной толщине мембраны.
Основная количественная характеристика, используемая при описании переноса ионов или незаряженных частиц (неэлектролитов) через мембраны- это поток.
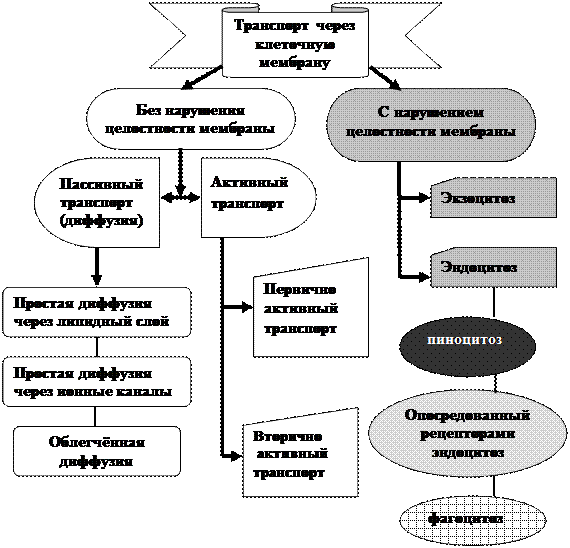 |
Рис.3.15. Виды транспорта веществ через клеточную мембрану
Поток вещества Ф измеряют не в числе частиц, а в числе молей данного вещества. Трансмембранные потоки ионов имеют направление нормальное к поверхности мембраны. Плотность потока (J, моль/с▪м2) – это количество вещества, переносимого в секунду через единицу площади, расположенной нормально потока (к поверхности мембраны). Положительным считается направление потока из замкнутого пространства наружу. Поэтому поток из клетки в окружающую среду имеет знак «+», а поток в клетку имеет знак «-». Величина J зависит от концентрации переносимых веществ по обе стороны мембраны (С1 и С2), а в случае ионов – так же от разности потенциалов между водными фазами, омывающими мембрану φ1 и φ2: ∆φ= φ2: - φ21. В конечном счёте обе характеристики (потенциал и концентрация) влияют на поток потому, что от них зависит энергия ионов (молекул) в данном растворе. Энергия моля ионов данного вида в среде носит название электрохимического потенциала иона и равна: μ=μо + RTlnC+zFφ, где μо – стандартный химический потенциал, определяемый энергией взаимодействия иона с молекулой среды;R- газовая постоянная; Т- абсолютная температура; С- молярная концентрация, z- безразмерный заряд иона, F- число Фарадея; φ – потенциал в данной области среды.
|
 |
Проницаемость клеточной мембраны для многих веществ ограничена не только липидным бислоем, но и прилегающими к мембране неперемешивающимися слоями воды, которые вместе с клеточной мембраной в совокупности ведут себя как последовательно соединённые электрические соединения. В свою очередь движение жидкости в клетке и вне её зависит от тканевого метаболизма. Если жидкость внутри клетки и в межклеточной среде интенсивно движется, то толщина примембранного слоя воды может превышать толщину самой клеточной мембраны в 100-300 раз. В то же время вязкость клеточной мембраны (благодаря липидам) в 10-100 раз выше вязкости воды. Следовательно, коэффициенты диффузии иона в воде и мембране относятся друг к другу как 30-100 к единице. Отсюда следует, что в коэффициенте распределения вещества в системе мембрана/вода решающую роль играет липофильность молекулы (иона). Если для К+, Na+, Cа2+ примембранные слои воды практически не влияют на общую проницаемость: основная барьерная функция принадлежит липидному бислою мембраны, то для нейтральных молекул (например кислорода) роль примембранных слоёв воды для его трансмембранного проникновения в клетку становится заметной.
Простая диффузия через ионные каналы мембраны (рис. 3.17.) даёт возможность проходить через клеточную мембрану веществам, обладающим низким коэффициентом распределения между липидной и водной фазами, следовательно, не растворяющихся в липидах. Например, вода, которая проходит сквозь мембрану клетки, благодаря наличию в ней специальных каналов (аквапоринов), расположенные в бислое липидов. При этом вода переходит из менее концентрированного раствора в более концентрированный. Через ионные каналы могут диффундировать малые незаряженные молекулы, в том числе, и способные одновременно проходить через липидный бислой (этанол, мочевина, О2, СО2 и др). Следует отметить избирательность ионных каналов для незаряженных катионов и анионов.
Ионные каналы клеточной мембраны представлены интегральными белками, которые образуют поры в липидном бислое клеточной мембран.
 |
Селективность (избирательность) канала определяется диаметром поры и энергией дегидратации иона. Диаметр поры несколько больше диаметра иона в кристаллическом состоянии. Малые размеры поры делают необходимым снятие с проходящего через мембрану иона его «водной шубы», т.е. осуществлять его временную дегидратацию.
Проницаемость канала регулируется а) изменением мембранного потенциала (потенциалуправляемые каналы); б) влиянием биологически активного вещества (гормоны, медиаторы, Са2+) на канальный белок (рецепторуправляемые каналы).
Для ионного канала характерны три физиологических состояния: покой (потенциальной активен), открыт (активирован) и закрыт (инактивирован).
|
 |
Рис. 3.18.
3. Наличие специфических стимуляторов и ингибиторов облегчённой диффузии. Например, инсулин активирует облегчённую диффузию глюкозы в жировые и мышечные клетки.
Активный транспорт
Данный вид переноса вещества через клеточную мембрану сопряжён с затратой энергии, благодаря чему он даёт возможность переносить вещества против градиента их концентрации, т.е. из зоны меньшей в зону большей концентрации. В качестве источника энергии в клетки используются: энергия связей аденозинтрифосфорной кислоты (АТФ) и энергия трансмембранных ионных градиентов. В зависимости от вида используемой энергии различают два вида активного транспорта: первично активный транспорт (используется энергия АТФ) и вторично активный транспорт (используется энергия электрохимического градиента Na+).
Первично активный транспорт осуществляется в результате деятельности ионных насосов, белковый комплекс которых обладает свойствами переносчика (для транспортируемого вещества) и фермента, расщепляющего АТФ, энергия которого используется насосом для транспорта (рис.3.19.).
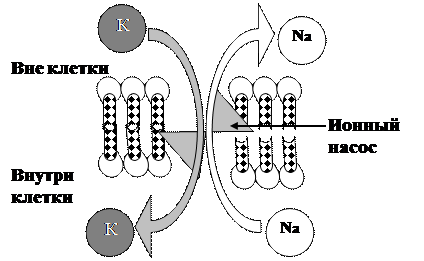
Рис.3.19. Схема первично активного транспорта через клеточную мембрану
В настоящее время в клетке обнаружены следующие насосы, использующие энергию АТФ: Na+,К+-насос (Na+,К+ -АТФаза) имеется в плазматической мембране всех клеток организма человека. Энергия АТФ затрачивается на удаление из клетки Na+ и возвращения туда К+, проникших путём простой диффузии. Са2+-насос (Са2+-АТФаза) как в плазматической мембране, так и мембранах клеточных органелл (например, цистерны в миоцитах). Насос откачивает кальций из цитозоля клетки либо во внеклеточную среду, либо в его внутриклеточные депо. Наличие данного насоса обусловлено той важной ролью, которую играет кальций в жизнедеятельности клеток. Протонный насос (Н+-АТФаза) имеется как в плазматической мембране, так и мембранах некоторых клеточных органелл.
Вторично активный транспорт (рис.3.20.) использует энергию градиента концентрации Na+ созданного К+,Na+-насосом. Ионы натрия и транспортируемые вещества связываются с молекулой белка переносчика мембраны. Натрий, перемещением внутрь клетки по электрохимическому градиенту «тащит» за собой вещества (например моносахара и аминокислоты), которые могут переносится против градиента концентрации. Внутриклеточный натрий откачивается Na+,К+- насосом, благодаря чему восстанавливается электрохимический градиент натрия и цикл повторяется снова.
 | |||||||
 |  | ||||||
 | |||||||





|

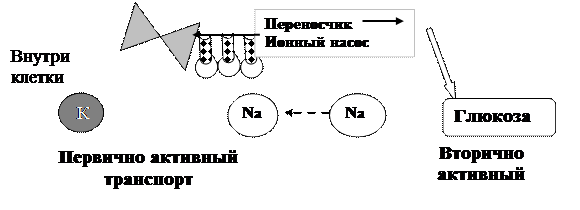 |
Рис. 3.20. Схема вторично активного транспорта через клеточную мембрану
Эндоцитоз и экзоцитоз
Данные виды трансмембранного переноса веществ в клетку и из неё сформировались в процессе эволюции и предназначены для крупномолекулярных веществ не способных перемещаться через клеточную мембрану разобранными выше способами.
Эндоцитоз заключается в поступлении вещества из внеклеточной среды в клетку в составе мелких или крупных пузырьков, образовавшихся из участков плазматической мембраны, в которых и заключено транспортируемое вещество. Эти пузырьки либо сливаются в клетках с органеллами (лизосомами), где происходит переваривание поступивших путём эндоцитоза веществ, либо они транспортируются на противоположный полюс клетки и выделяют воё содержимое во внеклеточную среду путём экзоцитоза.
Различают три вида эндоцитоза:
1. пиноцитоз ( неспецифических захват внеклеточной жидкости и растворённых в ней веществ)
2. опосредованный рецепторами эндоцитоз ( связывание веществ со специфическими рецепторами на клеточной мембране с образованием окаймлённых пузырьков (эндосом).
3. фагоцитоз (захват крупных частиц «профессиональными клетками» фагоцитами- нейтрофилы, моноциты, макрофаги)
Экзоцитоз – процесс выделения клеткой веществ во неклеточную среду. Он существует во всех клетках, но особенно он важен для нервных клеток (выделение медиаторов), клеток эндокринной системы (выделение гормонов) и клеток желёз внешней секреции (выделение пищеварительных соков). Выделение вещества из клетки происходит в составе экзоцитозных пузырьков, образующихся в специальной клеточной органелле (аппарат Гольджи). Пузырьки транспортируются с помощью микрофиламентов и микротрубочек к клеточной мембране, сливаются с ней, а их содержимое выделяется во внеклеточную среду.
Жизненный путь клетки
Клеточный цикл неполовых клеток состоит из интерфазы и митоза. Интерфаза включает в себя последовательно протекающие периоды G1, (G0)S, и G2. Период G1 следует за митозом. Длительность от нескольких часов до суток. После периода G1 наступает либо период G0, либо S. В период G0 клетка может выйти из цикла и либо: 1. Быть длительно неактивной (например, клетки иммунной памяти); 2.Функционировать как обычная дифференцированная клетка (например, гепатоцит) в обычных условиях; 3) Достигать необратимой дифференцировки (например, нейроны и кардиомиоцит ы). Если клетка переходит из периода G1 в период S (синтетический период), то через 8-12 часов переходит в премитотический период G2. который длится 2-4 часа и затем переходит в митоз. Митоз состоит из следующих фаз: профаза, метафаза, анафаза и телофаза. В ходе митоза из материнской клетки образуются две дочерние клетки, идентичные материнской.
Физиологическая гибель клетки (апопотоз) -это активный, генетически запрограммированный процесс. Пусковыми факторами апоптоза являются: нарушение рецепторного аппарата клетки, активные формы кислорода, этанол, низкокалорийную диету и т.д.. Существуют два пути его активации.
Первый путь реализуется через активацию специального гена, который запускает апоптоз путём остановки клеточного цикла и торможения ферментов, отвечающих за репарацию повреждённого участка ДНК. При этом актируются гены, кодирующие синтез белков-стимуляторов апопотоза, которые активируют ферменты, повреждающие структуры клетку и приводящие тем самым её к гибели. В клетке существуют и «гены-спасатели», кодирующие белки, тормозящие апоптоз.
Второй путь реализации апоптоза заключается в появлении на мембране стареющих клеток специального белка, к которому вырабатываются антитела, адсорбирующиеся на мембранах клеток с образованием комплекса белок-апоптоза + антитело. При его контакте с фагоцитом стареющая клетка, несущая данный комплекс, прикрепляется к мембране фагоцитирующей клетки и поглощается ею. Данный процесс получил название аутофагоцитоза.
Физиологическое значение апоптоза:
1) регуляция развития тканей и органов в период внутриутробного развития;
2) инволюция зрелых органов и тканей,
3)регуляция популяции клеток: уничтожения старых и генетически изменённых клеток в зрелых тканях.
Уменьшение способности к апоптозу ниже физиологического уровня –путь к аутоиммунным заболеваниям и злокачественному перерождению клеток.

