




Положение микробов среди живых существ.
| Эукариоты | Грибы Простейшие |
| Прокариоты | Актиномицеты Спирохеты Бактерии Микоплазмы Риккетсии Хламидии |
| Неклеточные | Вирусы |
Основные формы бактерий.
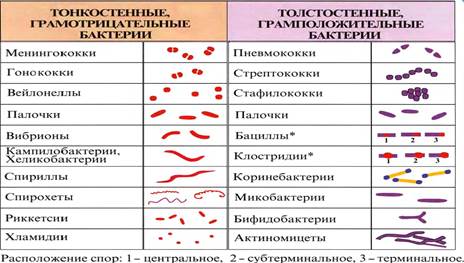
Бактерии бывают шаровидные, палочковидные, извитые и ветвящиеся. Кокки — шаровидные клетки, которые в зависимости от взаимного расположения делятся на микрококки, диплококки, стрептококки, тетракокки, сарцины, стафилококки.
Микрококки располагаются в виде отдельных клеток; диплококки, или парные кокки, — парами (пневмококк, гонококк, менингококк), так как клетки после деления не расходятся. Пневмококк (возбудитель пневмонии) имеет с противоположных сторон ланцетовидную форму, а гонококк (возбудитель гонореи) и менингококк (возбудитель эпидемического менингита) — форму кофейных зерен, обращенных вогнутой поверхностью друг к другу.
Стрептококки (от греч. streptos — цепочка) — клетки округлой или вытянутой формы, составляющие цепочку вследствие деления клеток в одной плоскости и сохранения связи между ними в месте деления. Сарцины (от лат. sarcina — связка, тюк) располагаются в виде пакетов из 8 и более кокков, так как они образуются при делении клетки в трех взаимно перпендикулярных областях. Стафилококки (от греч. staphyle — виноградная гроздь) представляют собой кокки, расположенные группами (гроздьями) в результате деления в разных плоскостях.
Палочковидные бактерии различаются по размерам, форме концов клетки и взаимному расположению клеток. Длина клеток варьирует от 1 до 8 мкм, толщина — от 0,5 до 2 мкм.
Палочки могут быть правильной (кишечная палочка и др.) и неправильной (коринебактерии и др.) формы, в том числе ветвящиеся, например актиномицеты. Наиболее мелкие палочковидные бактерии — риккетсии.
Концы палочек могут быть как бы обрезанными (сибиреязвенная бацилла), закругленными (кишечная палочка), заостренными (фузобактерии) или в виде утолщения, и тогда палочка похожа на булаву (коринебактерии дифтерии).
Слегка изогнутые палочки называют вибрионами (холерный вибрион). Большинство палочковидных бактерий располагается беспорядочно, так как после деления клетки расходятся.
Если после деления клетки остаются связанными общими фрагментами клеточной стенки и не расходятся, они располагаются под углом друг к другу (коринебактерии дифтерии), образуют цепочку (сибиреязвенная бацилла).
Извитые формы — спиралевидные бактерии, например спириллы, имеющие вид штопорообразно извитых клеток. К патогенным спириллам относится возбудитель содоку (болезни укуса крыс), к извитым — кампилобактеры, имеющие изгибы, как у крыла летящей чайки; близки к ним и такие бактерии, как спирохеты, имеющие ряд отличительных особенностей.
Структура бактериальной клетки. Постоянные и непостоянные структуры, их биологическая роль, способ выявления.
Структуру бактерий изучают с помощью электронной микроскопии целых клеток и их ультратонких срезов. Основными структурами бактериальной клетки являются: клеточная стенка, цитоплазматическая мембрана, цитоплазма с включениями и ядро, называемое нуклеоидом. Бактерии могут иметь и дополнительные структуры: капсулу, микрокапсулу, слизь, жгутики, фимбрии, пили; некоторые бактерии способны образовывать споры.
Размеры бактерий измеряют в микрометрах (мкм). Один микрометр равен 1000 нанометров (нм). В нанометрах измеряют отдельные компоненты бактерий.
Клеточная стенка — прочная, упругая структура, придающая бактерии определенную форму и сдерживающая высокое осмотическое давление в клетке. Она участвует в процессе деления клетки и транспорте метаболитов. У грамположительных бактерий клеточная стенка толще, чем у грамотрицательных, достигая 50 нм и более. В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов и белков. Большую часть массы D0—90 %) клеточной стенки этих бактерий составляет пептидогликан (синонимы: муреин,мукопептид), ковалентно связанный с тейхоевыми кислотами (от греч. teichos — стенка). В клеточной стенке грамотрицательных бактерий пептидогликана содержится меньше E—10 %).
Способность грамположительных бактерий при окраске по Граму удерживать генциановый фиолетовый в комплексе с йодом (сине-фиолетовая окраска бактерий) связана со свойством многослойного пептидогликана взаимодействовать с краской.
Обработка окрашенного по Граму мазка бактерий спиртом вызывает сужение пор в пептидогликане и тем самым задерживает краску в клеточной стенке. Наоборот, грамотрицательные бактерии после воздействия спиртом утрачивают краситель, обесцвечиваются и при обработке фуксином окрашиваются в красный цвет вследствие меньшего содержания пептидогликана E—10 % массы клеточной стенки).
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. Наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, называемой цитоплазматической мембраной. Основной компонент этих мембран —бимолекулярный (двойной) слой липидов. Наружная мембрана является асимметричной мозаичной структурой, представленной липополисахаридами, фосфолипидами и белками. С внешней стороны ее расположен липополисахарид (ЛПС), состоящий из трех компонентов: липида А, базисной части, или ядра (от лат. core — кор), и О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями. Липополисахарид «заякорен» в наружной мембране липидомА, придающим токсичность липополисахариду, отождествляемому поэтому с эндотоксином. От липида А отходит базисная часть липополисахарида. Наиболее постоянной частью ядра липополисахарида является кетодезоксиоктоновая кислота. О-специфическая цепь, отходящая от ядра липополисахарида, определяет серогруппу, серовар (разновидность бактерий, выявляемая с помощью иммунной сыворотки) определенного штамма бактерий. Таким образом, с понятием липополисахарида связаны представления об О-антигене, по которому можно дифференцировать бактерии.
Цитоплазма бактерий занимает основной объем клетки и состоит из растворимых белков. Рибосомы бактерий имеют коэффициент седиментации 70 S в отличие от рибосом, характерных для эукариотических клеток (80 S). Поэтому некоторые антибиотики, действие которых основано на подавлении синтеза белка путем связывания их с рибосомами бактерий, не оказывают влияния на синтез белка эукариотических клеток. В цитоплазме имеются различные включения — полисахариды, полимасляная кислота и полифосфаты (волютин). Они накапливаются при избытке питательных веществ в окружающей среде и выполняют роль запасных веществ для питания и энергетических потребностей. Зерна волютина выявляются у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки.
Нуклеоид (образование, подобное ядру) — эквивалент ядра у бактерий. Нуклеоид расположен в центральной зоне бактерий в виде двунитчатой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. В отличие от эукариот ядро бактерий не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК. При нарушении деления в ней может находиться 4 хромосомы и более.
Нуклеоид выявляется в световом микроскопе после окраски специфическими для ДНК методами по Фельгену или Гимзе. На электронограммах ультратонких срезов бактерий нуклеоид имеет вид светлых зон с фибриллярными, нитевидными структурами ДНК.
Кроме нуклеоида, представленного одной хромосомой, в бактериальной клетке имеются внехромосомные факторы наследственности — плазмиды.
Некоторые бактерии (пневмококки, клебсиеллы и др.) образуют капсулу — слизистое образование, прочно связанное с клеточной стенкой, имеющее четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже.
Она выявляется при специальных методах окраски, создающих негативное контрастирование вещества капсулы. Обычно капсула состоит из полисахаридов (экзополисахаридов), иногда полипептидов, например у сибиреязвенной бациллы. Капсула гидрофильна, она препятствует фагоцитозу бактерий. Многие бактерии образуют м икр о кап су л у — слизистое образование, выявляемое при электронной микроскопии. От капсулы следует отличать слизь — мукоидные экзополисахариды, не имеющие четких внешних границ. Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам), их еще называют гликокаликсом.
Жгутики бактерий определяют их подвижность. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны; длина их больше, чем длина клетки.
Толщина жгутиков 12—20 нм, длина — 3—12 мкм. Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих), у кишечной палочки, протея и др. ч.
Лофотрихи имеют пучок жгутиков на одном из концов клетки, амфитрихи — по одному жгутику или пучку жгутиков на противоположных концах клетки. Жгутики прикреплены к цитоплазматической мембране и клеточной стенке специальными дисками. По химическому составу жгутики состоят из белка — флагеллина (от англ. flagella — жгутик), обладающего антигенной специфичностью. Субъединицы флагеллина закручены в виде спирали. Жгутики выявляют с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами (рис. 2.4), или в световом микроскопе после обработки препаратов специальными методами (например, после серебрения).
Фимбрии и пили — нитевидные образования, более тонкие и короткие C—20 нм*0,3—10 мкм), чем жгутики.
Фимбрии отходят от поверхности клетки и состоят из белка, называемого пилином. Среди фимбрии разного типа выделяют фимбрии, ответственные за адгезию, т. е. прикрепление бактерий к поражаемой клетке (например, пили 3 общего типа — common pili); фимбрии, ответственные за питание, водно-солевой обмен; половые (F-пили), или конъюгационные, пили. Пили общего типа многочисленны и достигают нескольких сотен в одной клетке. Термин «пили» применяется чаще для обозначения особых фимбрий — половых пилей, образуемых так называемыми мужскими клетками-донорами, содержащими трансмиссивные плазмиды (F, R,Col); их количество 1—2 на клетку.
Отличительной особенностью половых пилей является взаимодействие с особыми «мужскими» сферическими бактериофагами.
Споры — своеобразная форма покоящихся фирмикутных бактерий, т. е. бактерий с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных условиях существования бактерий, сопровождающихся высушиванием, дефицитом питательных веществ и т. д. При этом внутри одной бактерии образуется одна спора. Поэтому образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие аэробные бактерии, у которых размер споры не превышает диаметра клетки, называются бациллами, а спорообразующие анаэробные бактерии, у которых размер споры превышает диаметр клетки и они поэтому принимают форму веретена, называются клостридиями (от лат. clostridium — веретено).

