




Электромагнитные явления в биологических системах.
Природа биопотенциалов и способы их описания.
Все процессы жизнедеятельности организмов сопровождаются появлением в клетках и тканях электродвижущих сил. Электрические явления играют большую роль в важнейших физиологических процессах: возбуждение клеток и проведение возбуждения по клеткам.
В возникновении биопотенциалов решающую роль играет разность потенциалов, обусловленная несимметричным распределением ионов. К таким разностям потенциалов относятся диффузионные, мембранные и фазовые.
1. Диффузионные потенциалы возникают на границе раздела двух жидких сред в результате различной подвижности ионов. Рассмотрим пример: Имеется раствор серной кислоты, разделенный пористой перегородкой. Пусть концентрация  в левой части больше, чем в пра
в левой части больше, чем в пра  вой. Ионы
вой. Ионы  и
и 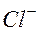 будут диффундировать из левой части в правую часть сосуда с разной скоростью по градиенту концентрации. Скорость диффузии определяется подвижностью ионов. Подвижность ионов больше подвижности ионов :
будут диффундировать из левой части в правую часть сосуда с разной скоростью по градиенту концентрации. Скорость диффузии определяется подвижностью ионов. Подвижность ионов больше подвижности ионов :  Следовательно, ионы будут намного опережать ионы . По обе стороны перегородки установятся потенциалы: слева “-“ справа “+”. Возникает диффузионная разность потенциалов. Эта разность потенциалов будет замедлять “быстрые“ ионы и ускорять “медленные“, т.е. возникающее электрическое поле направлено против сил диффузии. Диффузионная разность потенциалов максимален в тот момент, когда скорости диффузии становятся равными:
Следовательно, ионы будут намного опережать ионы . По обе стороны перегородки установятся потенциалы: слева “-“ справа “+”. Возникает диффузионная разность потенциалов. Эта разность потенциалов будет замедлять “быстрые“ ионы и ускорять “медленные“, т.е. возникающее электрическое поле направлено против сил диффузии. Диффузионная разность потенциалов максимален в тот момент, когда скорости диффузии становятся равными:
 .
.
Здесь 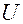 -;подвижность катионов;
-;подвижность катионов;  - подвижность анионов;
- подвижность анионов;  - универсальная газовая постоянная;
- универсальная газовая постоянная;  - абсолютная температура;
- абсолютная температура;  -валент-ность ионов;
-валент-ность ионов;  - число Фарадея;
- число Фарадея;  - активная концентрация в области откуда идет диффузия;
- активная концентрация в области откуда идет диффузия;  - активная концентрация в области куда идет диффузия.
- активная концентрация в области куда идет диффузия.
2. Мембранный потенциал. В рассматриваемом примере пористую перегородку заменим полупроницаемой мембраной, пропускающую только катионы – положительно заряженные ионы (это может быть мембрана с большой концентрацией фиксированных отрицательных ионов). 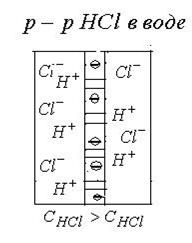
В этом случае подвижность ионов при переходе через мембрану не является определяющим фактором – в правую часть переходят только положительно заряженные ионы водорода. Через некоторое время диффузия ионов прекратится, т.к. они испытывают притяжение со стороны оставшихся в левой части ионов . Поэтому установится равновесие, в результате чего возникает двойной электрический слой: слева заряды “-“, справа заряды “+”, между сторонами мембраны возникнет разность потенциалов, называемая мембранным потенциалом, величина которого есть
 уравнение Нернста.
уравнение Нернста.
3. Фазовый потенциал возникает на границе раздела двух несмешивающихся фаз (например, раствор электролита в воде и какое-нибудь масло) в результате различной растворимости анионов и катионов в неводной фазе. Если, например, катионы растворимы в неводной фазе, то они активнее перейдут в нее и зарядят ее положительно относительно водной фазы. Наблюдается фазовый потенциал при механическом повреждении мембраны.
Равенство Доннана.
При выводе уравнений, описывающих распределение ионов между клеткой и окружающей средой, выполняется условие электронейтральности, равенство суммарной концентрации анионов (в основном и ионов макромолекул  ) катионов
) катионов 
как внутри клетки
 ,
,
так и снаружи клетки
 .
.
Здесь  -- число отрицательных зарядов на каждой белковой молекуле.
-- число отрицательных зарядов на каждой белковой молекуле.
В межклеточной жидкости содержание катионов значительно выше, чем ионов макромолекул . Поэтому
 - равенство Доннана
- равенство Доннана
Потенциал покоя
Экспериментально установлено, что цитоплазма в состоянии покоя имеет отрицательный потенциал, а окружающая среда- положительный.
Действительно, в первом приближении 
В клетке  в 20-40 раз.
в 20-40 раз.
Снаружи  в 10 раз.
в 10 раз.
Это неравномерное распределение концентраций обусловлено  насосом, при котором при переносе
насосом, при котором при переносе  переносится
переносится  . Кроме того, в клетке имеются анионы макромолекул (белков, аминокислот, и др.).
. Кроме того, в клетке имеются анионы макромолекул (белков, аминокислот, и др.).
Ионы внутри клетки не связаны с другими ионами и могут диффундировать туда, где их мало, т.е. в окружающую среду. Т.е. в состоянии покоя клетка проницаема только для ионов . Анионы не могут проникать через мембрану и остаются на внутренней поверхности мембраны. Т.о. мембрана снаружи зарядится положительно, а внутри – отрицательно.
Между внутренней и внешней поверхностями мембраны возникает разность потенциалов мембранной природы. Эта разность потенциалов между клеткой и окружающей средой, измеренная в состоянии физиологического покоя, называется потенциалом покоя. 
Если принять, в первом приближении, что потенциал покоя определяется только диффузией ионов , то величина потенциала покоя определяется как
 уравнение Нернста.
уравнение Нернста.
 - активная концентрация ионов внутри клетки,
- активная концентрация ионов внутри клетки,
 - активная концентрация ионов снаружи.
- активная концентрация ионов снаружи.
Если = , то  клетка мертва.
клетка мертва.
Но  , поэтому
, поэтому  . Например, для аксона гигантского кальмара
. Например, для аксона гигантского кальмара 
 , а это подтверждает, что в основе возникновения потенциала действия лежит перенос ионов.
, а это подтверждает, что в основе возникновения потенциала действия лежит перенос ионов.
В реальности в состоянии покоя мембрана проницаема не только для ионов , но и для ионов  и . Например, для аксона гигантского кальмара экспериментально установлено, что
и . Например, для аксона гигантского кальмара экспериментально установлено, что  . Основной вклад в потенциал покоя вносят ионы и . Ионов переносится очень мало. Поэтому на внешней поверхности сосредоточивается положительный заряд, а на внутренней – отрицательный. Потенциал покоя определяется тремя диффузионными потоками и вычисляется (с учетом равенства Доннана) по формуле
. Основной вклад в потенциал покоя вносят ионы и . Ионов переносится очень мало. Поэтому на внешней поверхности сосредоточивается положительный заряд, а на внутренней – отрицательный. Потенциал покоя определяется тремя диффузионными потоками и вычисляется (с учетом равенства Доннана) по формуле
 .
.
Кроме простой диффузии ионов и может идти обменная диффузия, но, как мы отмечали, потоки их равны и обменная диффузия не влияет на мембранный потенциал.
Потенциал действия.
Все клетки возбудимых тканей (нервная, мышечная, железистая) под действием различных раздражителей достаточной силы способны переходить в возбужденное состояние. Обязательным признаком возбуждения является изменение электрического состояния мембраны.
Опыт показывает, что возбужденный участок становится электроотри-цательным по отношению к невозбужденному участку. Следовательно, на возбужденном участке происходит перераспределение ионов. При возбуждении  это перераспределение кратковременно и концентрации восстанавливаются после снятия возбуждения, а разность потенциалов становится равной исходной, т.е. потенциалу покоя. Для аксона кальмара обнаружена такая зависимость изменения потенциала при возбуждении от времени
это перераспределение кратковременно и концентрации восстанавливаются после снятия возбуждения, а разность потенциалов становится равной исходной, т.е. потенциалу покоя. Для аксона кальмара обнаружена такая зависимость изменения потенциала при возбуждении от времени

 - потенциал покоя;
- потенциал покоя;  мембранный потенциал при возбуждении;
мембранный потенциал при возбуждении;
 - общее изменение разности потенциалов.
- общее изменение разности потенциалов.

