




В соответствии с функцией, выделяют следующие группы осязательных рецепторов: терморецепторы, механорецепторы и ноцирецепторы. Первые воспринимают изменения температуры, вторые – различные виды механических воздействий (прикосновение к коже, ее сдавливание), третьи – болевые раздражения.
Среди нервных окончаний различают свободные, лишенные глиальных клеток, и несвободные, у которых нервные окончания имеют оболочку – капсулу, образованную клетками нейроглии или соединительнотканными элементами.
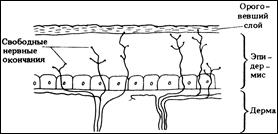
 Свободные нервные окончания
Свободные нервные окончания
имеются в коже. Они могут лежать в эпидермисе или прикрепляться к волоскам в волосяных фолликулах и реагируют на деформацию кожи или волоска. Рецепторы прикосновения сосредоточены в определенных участках тела и обусловливают повышенную чувствительность этих участков. Например, кончик языка может различать два стимула, разделенные расстоянием всего лишь 1 мм, тогда как в середине спины это возможно только в том случае, если расстояние между раздражаемыми точками составляет не менее 60 мм. Подходя к эпидермису, нервное волокно теряет миелин, проникает через базальную мембрану в эпителиальный слой, где разветвляется между эпителиоцитами вплоть до зернистого слоя. Конечные ветви диаметром менее 0,2 мкм на своих концах колбообразно расширяются. Аналогичные нервные окончания имеются в эпителии слизистых оболочек и в роговице глаза. Концевые свободные рецепторные нервные окончания являются ноцирецепторами (воспринимают боль) и терморецепторами (тепловыми и холодовыми).
Другие свободные нервные окончания проникают таким же образом в эпидермис и заканчиваются контактами с осязательными клетками (клетками Меркеля). Нервное окончание расширяется и образует с клеткой Меркеля синапсоподобный контакт. Эти окончания являются механорецепторами, воспринимающими давление.
Несвободные нервные окончания
могут быть инкапсулированными (покрыты соединительнотканной капсулой) и неинкапсулированными (лишены капсулы). Неинкапсулированные несвободные нервные окончания встречаются в соединительной ткани.
Инкапсулированными нервными окончаниями являются:
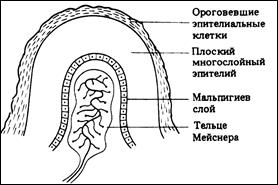 - тельца Фатера-Пачини (пластинчатые тельца) – самые крупные из всех инкапсулированных нервных окончаний. Они овальные, достигают 3-4 мм в длину и 2 мм в толщину. Располагаются в соединительной ткани внутренних органов и подкожной основе (дерме, чаще – на границе дермы и гиподермы). Большое число пластинчатых телец имеется в наружной оболочке крупных сосудов, в брюшине, сухожилиях и связках, по ходу шунтирующих сосудов. Тельце снаружи покрыто соединительнотканной капсулой, имеющей пластинчатое строение и богатой капиллярами. Под соединительнотканной оболочкой лежит наружная луковица, состоящая из 10-60 концентрических пластинок, образованных уплощенными клетками. Войдя в тельце, нервное волокно теряет миелиновую оболочку. Внутри тельца оно окружено лимфоцитами, которые формируют внутреннюю луковицу. Это механорецепторы, реагирующие прежде всего на вибрацию.
- тельца Фатера-Пачини (пластинчатые тельца) – самые крупные из всех инкапсулированных нервных окончаний. Они овальные, достигают 3-4 мм в длину и 2 мм в толщину. Располагаются в соединительной ткани внутренних органов и подкожной основе (дерме, чаще – на границе дермы и гиподермы). Большое число пластинчатых телец имеется в наружной оболочке крупных сосудов, в брюшине, сухожилиях и связках, по ходу шунтирующих сосудов. Тельце снаружи покрыто соединительнотканной капсулой, имеющей пластинчатое строение и богатой капиллярами. Под соединительнотканной оболочкой лежит наружная луковица, состоящая из 10-60 концентрических пластинок, образованных уплощенными клетками. Войдя в тельце, нервное волокно теряет миелиновую оболочку. Внутри тельца оно окружено лимфоцитами, которые формируют внутреннюю луковицу. Это механорецепторы, реагирующие прежде всего на вибрацию.
- тельца Мейснера (осязательные тельца) длиной 50-160 мкм и шириной около 60 мкм, овальные или цилиндрические. Их особенно много в коже пальцев. Они имеются также в коже губ, краев век, наружных половых органов. Тельце образовано множеством удлиненных, уплощенных или грушевидных лимфоцитов, лежащих один на другом. Нервные волокна, входящие в тельце, теряют миелин. Периневрий (соединительнотканная оболочка, покрывающая пучок нервных волокон) переходит в окружающую тельце капсулу, образованную несколькими слоями клеток. Тельца Мейснера являются механорецепторами, воспринимающими прикосновение и сдавливание кожи.
- тельца Гольджи-Маццони – луковицеобразные механорецепторы.
- тельца Руффини – веретенообразные, расположены в коже пальцев кисти и стопы, в капсулах суставов и стенках кровеносных сосудов. Тельце окружено тонкой капсулой, образованной клетками периневрия. Войдя в капсулу, нервное волокно теряет миелин и разветвляется на множество ветвей, которые заканчиваются колбообразными вздутиями, окруженными микроглиальными клетками. Окончания плотно прилегают к фибробластам и коллагеновым волокнам, формирующим основу тельца. Тельца Руффини являются механорецепторами, они также воспринимают тепло и служат проприорецепторами.
- колбы Краузе (концевые колбы) сферические по форме, расположены в коже, конъюнктиве глаз, слизистой оболочке рта. Колба имеет толстую соединительнотканную капсулу. Войдя в капсулу, нервное волокно теряет миелиновую оболочку и разветвляется в центре колбы, образуя множество ветвей. Колбы Краузе воспринимают холод; возможно, они являются и механорецепторами.
Хеморецепция.
Ощущения вкуса и запаха связаны с действием химических веществ, возбуждающих специфические хеморецепторы и играющих важную роль в питании, избегании опасности и других поведенческих реакциях, включая ухаживание, спаривание и социальную активность.
Вкус
Многие ощущения, воспринимаемые как вкусовые, на самом деле обусловлены раздражением как вкусовых, так и обонятельных рецепторов. Если человек с завязанными глазами и зажатым носом (чтобы химические стимулы не действовали на орган обоняния) будет жевать лук или яблоко, вряд ли он сможет их различить. Это свидетельствует о малой чувствительности вкусовых рецепторов. Молекулы вкусовых раздражителей взаимодействуют со специфическими рецепторными белками, находящимися в плазмалемме микроворсинок сенсорных клеток. Эти клетки погруженны в бокаловидные образования – вкусовые почки – на верхней и боковых поверхностях языка (рис. А).
Существуют четыре типа вкусовых ощущений, и неравномерное распределение воспринимающих их рецепторов можно продемонстрировать, нанося на разные участки языка различные вещества: сладкое (сахарин), кислое (слабую кислоту), соленое (морскую соль) и горькое (хинин). Распределение соответствующих рецепторных зон показано на рис. Б. Рецепторный потенциал возникает в рецепторной клетке под действием веществ, проникающих через ее мембрану или взаимодействующих с рецепторными молекулами на поверхности этой мембраны. Для стимуляции вкусовых рецепторов требуются более высокие концентрации веществ, чем для стимуляции обонятельных рецепторов
А. Схема строения вкусовой почки. Обладающие вкусом вещества растворяются в жидкости, омывающей микроворсинки, и диффундируют в рецепторные клетки.
Б. Симметричное распределение вкусовых рецепторов и их иннервация в языке человека.
Обоняние
 Обонятельные ощущения обладают большей остротой, чем вкусовые, и вызываются пахучими веществами, находящимися в воздухе. Эти вещества растворяются в слизи и стимулируют обонятельный эпителий, расположенный в верхней части носовой полости. В каждой половине носа имеется обонятельная область площадью около 2,5 см2 с клетками трех типов - сенсорными, опорными и базальными (см. рис.).
Обонятельные ощущения обладают большей остротой, чем вкусовые, и вызываются пахучими веществами, находящимися в воздухе. Эти вещества растворяются в слизи и стимулируют обонятельный эпителий, расположенный в верхней части носовой полости. В каждой половине носа имеется обонятельная область площадью около 2,5 см2 с клетками трех типов - сенсорными, опорными и базальными (см. рис.).
В 1963 г. Аморе (Amoore) выдвинул стереохимическую теорию обоняния, согласно которой существует соответствие между свойствами рецепторных молекул, находящихся в ресничках обонятельных клеток, и конфигурацией и размерами молекул пахучих веществ. Он предположил, что существует семь основных запахов, таких как гнилостный, острый, едкий и т.д., каждому из которых соответствуют свои специфические рецепторы. Всего можно различать около 10 000 запахов, что, по мнению Аморе, связано с разной степенью стимуляции рецепторов тех или иных типов. Наиболее сильные обонятельные ощущения вызывают вещества, растворимые в воде и жирах.
Упрощенная схема взаимоотношений между сенсорными, опорными и базальными клетками в обонятельном эпителии. Молекулы-рецепторы для пахучих веществ находятся на поверхностных мембранах ресничек обонятельного эпителия.

Электрорецепторы (дополнительный материал)
В коже некоторых костистых и хрящевых рыб волосковые клетки утратили волоски и превратились в детекторы электрических токов в воде. Источниками токов служат или сами эти рыбы (электрические органы, генерирующие низковольтные разряды, имеются, например, у представителей семейства Клюворылые – Mormyridae), или электрически активные ткани других животных, находящихся поблизости. Способность определять локализацию добычи по токам, исходящим от ее работающих мышц, особенно развита у некоторых акул.
Электрорецепторы у рыб находятся на голове и теле в системе боковой линии. У «низковольтных» электрических рыб (в отличие от «высоковольтных» – электрический угорь) электрические разряды, генерируемые на одном конце тела, входят обратно в тело рыбы через эпителиальные поры в боко  вой линии. Разряды генерируются видоизмененной мышечной или нервной тканью
вой линии. Разряды генерируются видоизмененной мышечной или нервной тканью
У основания каждой поры ток встречает на своем пути электрорецепторную клетку (см. рисунок справа), образующую синаптические контакты с терминалями VIII черепномозгового нерва, иннервирующими боковую линию. Апикальная мембрана рецепторной клетки (обращенная к внешней среде) обладает меньшим электрическим сопротивлением, чем мембрана на базальной стороне клетки. Поэтому наибольшее падение потенциала происходит в месте выхода тока из клетки. Таким образом, ток, входящий из окружающей среды в пору и проходящий через электрорецепторную клетку, вызывает деполяризацию мембраны в базальной области. Деполяризация активирует кальциевые каналы в этой части клеточной мембраны, и приток Ca2+ в клетку через эти каналы ускоряет освобождение синаптического медиатора, а это в свою очередь ведет к повышению частоты импульсов в сенсорном волокне, иннервирующем рецептор. Наоборот, ток, выходящий из тела рыбы, гиперполяризует мембрану в основании рецепторной клетки и уменьшает выделение медиатора ниже спонтанного уровня. Таким образом, частота импульсов в аксонах может уменьшаться или увеличиваться в зависимости от направления тока, протекающего через электрорецепторную клетку. Подобно волосковым клеткам уха позвоночных, эти рецепторы и иннервирующие их терминали обладают поистине удивительной чувствительностью к внешним стимулам: частота разрядов в сенсорном нерве изменяется даже при сдвиге мембранного потенциала рецепторной клетки, составляющем всего лишь несколько микровольт (миллионных долей вольта).
Некоторые угри, скаты и другие рыбы генерируют мощный разряд электрического тока, которым они оглушают врага или добычу. В отличие от них «низковольтные» электрические рыбы генерируют своими электрическими органами непрерывные серии синхронных деполяризации довольно высокой частоты. Серии электрических импульсов распространяются в воде от заднего конца тела к переднему. Любой объект, электропроводность которого отличается от электропроводности воды, вызывает искажение линий тока. Электрорецепторы боковой линии «следят» за распределением тока, возвращающегося через поры боковой линии на голове и переднем конце тела, и способны выявлять изменения электрического поля, обусловленные присутствием в воде посторонних объектов. Эта сенсорная информация перерабатывается затем в имеющем очень крупные относительные размеры мозжечке рыбы, что позволяет ей обнаруживать в непосредственной от себя близости объекты и определять их местонахождение.
Орган слуха и равновесия (преддверно-улитковый орган)

