




Главные ферменты, контролирующие метаболизм гликогена, - гликогенфосфорилаза и гликогенсинтаза – регулируются аллостерически и ковалентной модификацией путем фосфорилирования и дефосфорилирования фермента.
1. В печени фосфорилаза находится в активной и неактивной форме. Активная фосфорилаза (фосфорилаза а) – фосфорилированный тетрамер. Под действием специфической фосфатазы фосфорилаза а переходит в неактивную фосфорилазу b (димер) в результате гидролитического отщепления фосфата от остатка серина. Переход неактивной фосфорилазы b в фосфорилазу а происходит путем фосфорилирования под действием киназы фосфорилазы.
Соотношение между неактивным Т-состоянием фосфорилазы b и активным R-состоянием фосфорилазы а определяется энергетическим зарядом мышечной клетки. Глюкозо-6-фосфат и АТФ стабилизируют неактивное Т-состояние фермента (фосфорилаза b). Фосфорилаза а всегда активна, независимо от концентраций АМФ, АТФ и глюкозо-6-фосфата. В покое преобладает фосфорилаза b, а при работе АМФ переводит фермент в активную форму а.
2. Киназа фосфорилазы существует в 2-х формах – активной – фосфорилированной и неактивной – дефосфорилированной. Переход активной формы в неактивную осуществляется при отщеплении фосфорной кислоты. Активация киназы фосфорилазы происходит при участии фермента протеинкиназы.
3. Гликогенсинтаза может находиться в фосфорилированном и нефосфорилированном состоянии. Активна дефосфорилированная форма фермента – гликогенсинтаза а, которая переходит в неактивную гликогенсинтазу b (зависима от концентрации глюкозо-6-фосфата) путем фосфорилирования гидроксильных групп остатков серина под действием протеинкиназы. Превращение гликогенсинтазы b в активную форму а происходит под действием протеинфосфатазы путем отщепления фосфатных групп от остатков серина.
4. Протеинкиназа состоит из 4-х субъединиц (тетрамер) – 2-х регуляторных (R) и 2-х каталитических (С). В комплексе R2C2 фермент неактивен и активируется под действием циклического АМФ (цАМФ). 2 молекулы цАМФ присоединяются к каждой регуляторной субъединице протеинкиназы, что вызывает его диссоциацию с высвобождением каталитических субъединиц.
R2C2 + 4цАМФ → 2 С + 2 (R – 2 цАМФ)
Активная протеинкиназа переносит остаток фосфорной кислоты от АТФ на специфические белки (ферменты), что приводит к изменению их активности.
5. Образование цАМФ (3′5′-АМФ) происходит из АТФ под действием фермента аденилатциклазы, который находится на внутренней поверхности мембраны и связан с рецептором для гормонов на внешней стороне мембраны.
6. Фермент фосфодиэстераза разрушает цАМФ путем разрыва кольца с образованием АМФ.
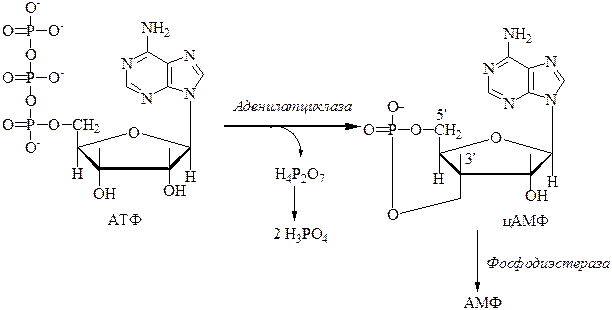
При голодании или в стрессовых ситуациях увеличивается потребность в глюкозе.
1. В мышце под действием адреналина, а в печени – глюкагона и адреналина происходит активация аденилатциклазы, которая приводит к образованию цАМФ, который, связываясь с R-субъединицей протеинкиназы, активирует ее. Протеинкиназа фосфорилирует киназу фосфорилазу b и переводит ее в активную форму (киназу фосфорилазы а). Киназа фосфорилазы a фосфорилирует фосфорилазу b и переводит ее в фосфорилазу а, что активирует распад гликогена. Инсулин ингибирует аденилатциклазу и активирует фосфодиэстеразу, что уменьшает концентрацию цАМФ. Протеинкиназа, киназа фосфорилазы и фосфорилаза остаются в неактивной форме, что тормозит распад гликогена. Таким образом, происходит каскад ферментативных реакций, что создает высокую степень амплификации. При распаде гликогена – 3 ферментативных стадии контроля, при синтезе – 2.
2. В мышцах фосфорилаза b может активироваться аллостерически под действием АМФ, оставаясь при этом дефосфорилированной. АТФ действует как отрицательный аллостерический эффектор.
3. В мышцах сразу после мышечного сокращения гликогенолиз возрастает в несколько сотен раз. Процесс включает быструю активацию фосфорилазы через ионы кальция. Ионы Са2+ активируют киназу фосфорилазы, связываясь с субъединицей киназы фосфорилазы b, идентичной Са2+-связывающему белку – кальмодулину.
4. Гликогенсинтаза b может активироваться аллостерически под действием глюкозо-6-фосфата. Инсулин стимулирует синтез гликогена в мышце, способствуя дефосфорилированию и активации гликогенсинтазы.
 Рис. 4. Регуляция синтеза и распада гликогена
Рис. 4. Регуляция синтеза и распада гликогена
Регуляция распада гликогена носит каскадный характер.

