




Аэробный распад глюкозы
Энергетическое значение аэробного распада глюкозы.
В аэробном гликолизе образуется 10 моль АТФР на 1 моль глюкозы. Так, в реакциях 7, 10 образуется 4 моль АТФ путем субстратного фосфорилирования, а в реакции 6 синтезируется 6 моль АТФ (на 2 моль глицероальдегидфосфата) путем окислительного фосфорилирования.
Баланс аэробного гликолиза.
Суммарный эффект аэробного гликолиза составляет 8 моль АТФ, так как в реакциях 1 и 3 используется 2 моль АТФ. Дальнейшее окисление двух моль пируват в общих путях катаболизма сопровождается синтезом 30 моль АТФ (по 15 моль на каждую молекулу пирувата. Следовательно, суммарный энергетический эффект аэробного распада глюкозы до конечных продуктов составляет 38 моль АТФ.
Значение анаэробного гликолиза
Анаэробный и аэробный гликолиз энергетически неравноценны. Образование двух моль лактата из глюкозы сопровождается синтезом всего двух моль АТФ, потому что NADH, полученный при окислении глицероальдегидфосфата, не используется дыхательной цепью, а акцептируется пируватом.
Анаэробный распад глюкозы.
Анаэробный гликолиз, несмотря на небольшой энергетический эффект, является основным источником энергии для скелетных мышц в начальном периоде интенсивной работы, т. е. в условиях, когда снабжение кислородом ограничено. Кроме того, зрелые эритроциты извлекают энергию за счет анаэробного окисления глюкозы, потому что не имеют митохондрий.
Спиртовое брожение — химическая реакция брожения, осуществляемая дрожжами, в результате которой одна молекула глюкозы преобразуется в 2 молекулы этанола и в 2 молекулы углекислого газа.
Спиртовое брожение (осуществляется дрожжами и некоторыми видами бактерий), в ходе него пируват расщепляется на этанол и двуокись углерода. Из одной молекулы глюкозы в результате получается две молекулы питьевого спирта (этанола) и две молекулы углекислого газа. Этот вид брожения очень важен в производстве хлеба, пивоварении, виноделии и винокурении. Если в закваске высока концентрация пектина, может также производиться небольшое количество метанола. Обычно используется только один из продуктов; в производстве хлеба алкоголь улетучивается при выпечке, а в производстве алкоголя двуокись углерода обычно уходит в атмосферу, хотя в последнее время её стараются утилизировать.
40.Глюконеогенез — процесс образования в печени и отчасти в корковом веществе почек (около 10 %) молекул глюкозы из молекул других органических соединений — источников энергии, например свободных аминокислот, молочной кислоты,глицерина
.Суммарное уравнение глюконеогенеза: 2 CH3COCOOH + 4ATP + 2GTP + 2NADH.H+ + 6 H2O = C6H12O6 + 2NAD + 4ADP + 2GDP + 6Pn [1] .
Роль в организме
При голодании в организме человека активно используются запасы питательных веществ (гликоген, жирные кислоты). Они расщепляются до аминокислот, кетокислот и других неуглеводных соединений. Большая часть этих соединений не выводится из организма, а подвергаются реутилизации. Вещества транспортируются кровью в печень из других тканей, и используются в глюконеогенезе для синтеза глюкозы — основного источника энергии в организме. Таким образом при истощении запасов организма, глюконеогенез является основным поставщиком энергетических субстратов.
Большинство стадий глюконеогенеза представляет собой обращениереакциигликолиза. Только 3 реакциигликолиза (гексокиназная, фосфо-фруктокиназная и пируваткиназная) необратимы, поэтому в процесс глю-конеогенеза на 3 этапах используются другие ферменты. Рассмотрим путь синтеза глюкозы из пирувата.
Образование фосфоенолпирувата из пирувата. Синтез фосфоенолпирувата осуществляется в несколько этапов. Первоначально пируват под влиянием
пируваткарбоксилазы и при участии СО2 и АТФ карбоксилируется с образованием оксалоацетата:
 Затем оксалоацетат в результате декарбоксилирования и фосфорили-рования под влиянием фермента
фосфоенолпируваткарбоксилазы превращается в фосфоенолпируват. Донором фосфатного остатка в реакции
служит гуанозинтрифосфат (ГТФ):
Затем оксалоацетат в результате декарбоксилирования и фосфорили-рования под влиянием фермента
фосфоенолпируваткарбоксилазы превращается в фосфоенолпируват. Донором фосфатного остатка в реакции
служит гуанозинтрифосфат (ГТФ):
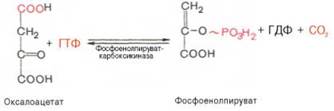 Установлено, что в процессе образования фосфоенолпирувата участвуют ферменты цитозоля и митохондрий.
Первый этап синтеза протекает в митохондриях (рис. 10.6). Пируват-карбоксилаза, которая катализирует эту реакцию, является аллостери-ческим митохондриальным ферментом. В качестве аллостерического активатора данного фермента необходим ацетил-КоА. Мембранамитохондрий непроницаема для образовавшегося оксалоацетата. Последний здесь же, в митохондриях, восстанавливается в малат:
Установлено, что в процессе образования фосфоенолпирувата участвуют ферменты цитозоля и митохондрий.
Первый этап синтеза протекает в митохондриях (рис. 10.6). Пируват-карбоксилаза, которая катализирует эту реакцию, является аллостери-ческим митохондриальным ферментом. В качестве аллостерического активатора данного фермента необходим ацетил-КоА. Мембранамитохондрий непроницаема для образовавшегося оксалоацетата. Последний здесь же, в митохондриях, восстанавливается в малат:
 Реакция протекает при участии митохондриальной НАД-зависимой малатдегидрогеназы. В митохондриях отношение НАДН/НАД+ относительно велико, в связи с чем внутримитохондриальный оксалоацетат легко восстанавливается в малат, который легко выходит из митохондрии через митохондриальную мембрану. В цитозоле отношение НАДН/НАД+ очень мало, и малат вновь окисляется при участии цитоплазматической НАД-за-висимой малатдегидрогеназы:
Реакция протекает при участии митохондриальной НАД-зависимой малатдегидрогеназы. В митохондриях отношение НАДН/НАД+ относительно велико, в связи с чем внутримитохондриальный оксалоацетат легко восстанавливается в малат, который легко выходит из митохондрии через митохондриальную мембрану. В цитозоле отношение НАДН/НАД+ очень мало, и малат вновь окисляется при участии цитоплазматической НАД-за-висимой малатдегидрогеназы:
 Дальнейшее превращение оксалоацетата в фосфоенолпируват происходит в цитозоле клетки.
Превращение фруктозо-1,6-бисфосфата во фруктозо-6-фосфат. Фосфо-енолпируват, образовавшийся из пирувата, в результате ряда обратимых реакцийгликолиза превращается во фруктозо-1,6-бисфосфат. Далее следует фосфофруктокиназная реакция, которая необратима. Глюконеогенез идет в обход этой эндергонической реакции. Превращение фруктозо-1,6-бис-фосфата во фруктозо-6-фосфат катализируется специфической фосфатазой:
Дальнейшее превращение оксалоацетата в фосфоенолпируват происходит в цитозоле клетки.
Превращение фруктозо-1,6-бисфосфата во фруктозо-6-фосфат. Фосфо-енолпируват, образовавшийся из пирувата, в результате ряда обратимых реакцийгликолиза превращается во фруктозо-1,6-бисфосфат. Далее следует фосфофруктокиназная реакция, которая необратима. Глюконеогенез идет в обход этой эндергонической реакции. Превращение фруктозо-1,6-бис-фосфата во фруктозо-6-фосфат катализируется специфической фосфатазой:
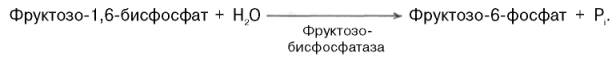 .Образование глюкозы из глюкозо-6-фосфата. В последующей обратимой стадии биосинтезаглюкозы фруктозо-6-фосфат превращается в глюкозо-6-фосфат. Последний может дефосфорилироваться (т.е. реакция идет в обход гексокиназной реакции) под влиянием фермента глюкозо-6-фос-фатазы:
.Образование глюкозы из глюкозо-6-фосфата. В последующей обратимой стадии биосинтезаглюкозы фруктозо-6-фосфат превращается в глюкозо-6-фосфат. Последний может дефосфорилироваться (т.е. реакция идет в обход гексокиназной реакции) под влиянием фермента глюкозо-6-фос-фатазы:
 Регуляция глюконеогенеза. Важным моментом в регуляции глюконеоге-неза является реакция, катализируемая пируваткарбоксилазой. Роль положительного аллостерического модулятора этого фермента выполняет ацетил-КоА. В отсутствие ацетил-КоА фермент почти полностью лишен активности. Когда в клетке накапливается митохондриальный ацетил-КоА, биосинтезглюкозы из пирувата усиливается. Известно, что ацетил-КоА одновременно является отрицательным модулятором пируватдегидроге-назного комплекса (см. далее). Следовательно, накопление ацетил-КоА замедляет окислительное декарбоксилирование пирувата, что также способствует превращению последнего в глюкозу.
Другой важный момент в регуляции глюконеогенеза – реакция, катализируемая фруктозо-1,6-бисфосфатазой – ферментом, который ингибиру-ется АМФ. Противоположное действие АМФ оказывает на фосфофруктокиназу, т. е. для этого фермента он является аллостерическим активатором. При низкой концентрацииАМФ и высоком уровне АТФ происходит стимуляция глюконеогенеза. Напротив, когда величина отношения АТФ / АМФ мала, в клетке наблюдается расщепление глюкозы.
Показано, что глюконеогенез может регулироваться и непрямым путем, т.е. через изменение активности фермента, непосредственно не участвующего в синтезе глюкозы. Так, установлено, что ферментгликолиза пиру-ваткиназа существует в 2 формах – L и М. Форма L (от англ. liver – печень) преобладает в тканях, способных к глюконеогенезу. Эта форма ингиби-руется избытком АТФ и некоторыми аминокислотами, в частности ала-нином. М-форма (от англ. muscle – мышцы) такой регуляции не подвержена. В условиях достаточного обеспечения клетки энергией происходит инги-бирование L-формы пируваткиназы. Как следствие ингибирования замедляется гликолиз и создаются условия, благоприятствующие глюконеоге-незу.
Наконец, интересно отметить, что между гликолизом, интенсивно протекающим в мышечной ткани при ее активной деятельности, и глюко-неогенезом, особенно характерным для печеночной ткани, существует тесная взаимосвязь. При максимальной активности мышц в результате усиления гликолиза образуется избыток молочной кислоты, диффундирующей в кровь, в печени значительная ее часть превращается в глюкозу (глюконеогенез). Такая глюкоза затем может быть использована как энергетический субстрат, необходимый для деятельности мышечной ткани.
Регуляция глюконеогенеза. Важным моментом в регуляции глюконеоге-неза является реакция, катализируемая пируваткарбоксилазой. Роль положительного аллостерического модулятора этого фермента выполняет ацетил-КоА. В отсутствие ацетил-КоА фермент почти полностью лишен активности. Когда в клетке накапливается митохондриальный ацетил-КоА, биосинтезглюкозы из пирувата усиливается. Известно, что ацетил-КоА одновременно является отрицательным модулятором пируватдегидроге-назного комплекса (см. далее). Следовательно, накопление ацетил-КоА замедляет окислительное декарбоксилирование пирувата, что также способствует превращению последнего в глюкозу.
Другой важный момент в регуляции глюконеогенеза – реакция, катализируемая фруктозо-1,6-бисфосфатазой – ферментом, который ингибиру-ется АМФ. Противоположное действие АМФ оказывает на фосфофруктокиназу, т. е. для этого фермента он является аллостерическим активатором. При низкой концентрацииАМФ и высоком уровне АТФ происходит стимуляция глюконеогенеза. Напротив, когда величина отношения АТФ / АМФ мала, в клетке наблюдается расщепление глюкозы.
Показано, что глюконеогенез может регулироваться и непрямым путем, т.е. через изменение активности фермента, непосредственно не участвующего в синтезе глюкозы. Так, установлено, что ферментгликолиза пиру-ваткиназа существует в 2 формах – L и М. Форма L (от англ. liver – печень) преобладает в тканях, способных к глюконеогенезу. Эта форма ингиби-руется избытком АТФ и некоторыми аминокислотами, в частности ала-нином. М-форма (от англ. muscle – мышцы) такой регуляции не подвержена. В условиях достаточного обеспечения клетки энергией происходит инги-бирование L-формы пируваткиназы. Как следствие ингибирования замедляется гликолиз и создаются условия, благоприятствующие глюконеоге-незу.
Наконец, интересно отметить, что между гликолизом, интенсивно протекающим в мышечной ткани при ее активной деятельности, и глюко-неогенезом, особенно характерным для печеночной ткани, существует тесная взаимосвязь. При максимальной активности мышц в результате усиления гликолиза образуется избыток молочной кислоты, диффундирующей в кровь, в печени значительная ее часть превращается в глюкозу (глюконеогенез). Такая глюкоза затем может быть использована как энергетический субстрат, необходимый для деятельности мышечной ткани.
|
41. Гликоген - основная форма депонирования глюкозы в клетках животных. У растений эту же функцию выполняет крахмал. В структурном отношении гликоген, как и крахмал, представляет собой разветвленный полимер из глюкозы.
Однако гликоген более разветвлен и компактен. Ветвление обеспечивает быстрое освобождение при распаде гликогена большого количества концевых мономеров. Синтез и распад гликогена не являются обращением друг в друга, эти процессы происходят разными путями.
Биосинтез***гликогена
Гликоген синтезируется в период пищеварения (в течение 1-2 ч после приема углеводной пищи). Гликогенез особенно интенсивно протекает в печени и скелетных мышцах. В начальных реакциях образуется UDF-глюкоза (реакция 3), которая является активированной формой глюкозы, непосредственно включающейся в реакцию полимеризации (реакция 4). Эта последняя реакция катализируется гликогенсинтазой, которая присоединяет глюкозу к олигосахариду или к уже имеющейся в клетке молекуле гликогена, наращивая цепь новыми мономерами. Для подготовки и включения в растущую полисахаридную цепь требуется энергия 1 моль АТР и 1 моль UTP. Ветвление полисахаридной цепи происходит при участии фермента амило - -1,4--1,6-гликозил-трансферазы путем разрыва одной -1,4-связи и переноса олигосахаридного остатка от конца растущей цепи к ее середине с образованием в этом месте -1,6-гликозидной связи. Молекула гликогена содержит до 1 млн остатков глюкозы, следовательно, на синтез расходуется значительное количество энергии. Необходимость превращения глюкозы в гликоген связана с тем, что накопление значительного количества глюкозы в клетке привело бы к повышению осмотического давления, так как глюкоза хорошо растворимое вещество. Напротив, гликоген содержится в клетке в виде гранул, и мало растворим. Распад гликогена - гликогенолиз - происходит в период между приемами пищи.
Второй вариант билета 40.
Биосинтез глюкозы - глюконеогенез
Глюконеогенез - это синтез глюкозы из неуглеводных предшественников. У млекопитающих эту функцию выполняет в основном печень, в меньшей мере - почки и клетки слизистой кишечника. Запасов гликогена в организме достаточно для удовлетворения потребностей в глюкозе в период между приемами пищи. При углеводном или полном голодании, а также в условиях длительной физической работы концентрация глюкозы в крови поддерживается за счет глюконеогенеза. В этот процесс могут быть вовлечены вещества, которые способны превратиться в пируват или любой другой метаболит глюконеогенеза.
Причем, использование первичных субстратов в глюконеогенезе происходит в различных физиологических состояниях. Так, в условиях голодания часть тканевых белков распадается до аминокислот, которые затем используются в глюконеогенезе. При распаде жиров образуется глицерин, который через диоксиацетонфосфат включается в глюконеогенез. Лактат, образующийся при интенсивной физической работе в мышцах, затем в печени превращается в глюкозу. Следовательно, физиологическая роль глюконеогенеза из лактата и из аминокислот и глицерина различна. Синтез глюкозы из пирувата протекает, как и при гликолизе, но в обратном направлении.
Глюконеогенез.
Ферменты: 1-пируваткарбоксилаза, 2-фосфоенолпируваткарбоксикиназа, 3-фосфатаза фру-1,6-дифосфата, 4-глюкозо-6-фосфатаза.
Семь реакций гликолиза легко обратимы и используются в глюконеогенезе. Но три киназные реакции необратимы и должны шунтироваться. Так, фруктозо-1,6-дифосфат и глюкозо-6-фосфат дефосфорилируются специфическими фосфатазами, а пируват фосфорилируется до образования фосфоенолпирувата посредством двух промежуточных стадий через оксалоацетат. Образование оксалоацетата катализируется пируваткарбоксилазой. Этот фермент содержит в качестве кофермента биотин. Оксалоацетат образуется в митохондриях, транспортируется в цитозоль и включается в глюконеогенез. Следует обратить внимание на то, что каждая из необратимых реакций гликолиза вместе с соответствующей ей необратимой реакцией глюконеогенеза составляют цикл, называемый субстратным.
Глюконеогенез, необратимые реакции.
Таких циклов существует три - соответственно трем необратимым реакциям. Результатом одновременного протекания реакций субстратных циклов будет расходование энергии. Субстратные циклы могут протекать в условиях нормального обмена веществ в печени и имеют вполне определенное биологическое значение. Кроме того, эти циклы служат точками приложения регуляторных механизмов, в результате чего изменяется поток метаболитов либо по пути распада глюкозы, либо по пути ее синтеза. Суммарное уравнение глюконеогенеза из пирувата:
2 пируват + 4 ATФ + 2 GTP + 2(NADH) + 4 Н2О Глюкоза + 4 ADP + 2 GDP + 2 NAD+ + 6 Н3РО4.
За сутки в организме человека может синтезироваться до 80 г глюкозы. На синтез 1 моль глюкозы из пирувата расходуется 6 макроэргических связей (4 ATФ и 2 GTP).

