




Где были обнаружены свойства, которыми не обладает зрительная кора (целостность и константность): заднетеменная и нижневисочная кора.
Часть волокон, идущих от сетчатки, идёт к структуре, которая называется верхнее двухолмие. Это слоистая структура (6 слоев), в которой расположены нейроны, обеспечивающие управление саккадическими движениями глаз (перевод взгляда с одного объекта на другой). Информация от клеток верхнего двухолмия идет в заднетеменную кору.
В заднетеменной коре расположены нейроны, которые имеют рецептивные поля и связаны не с изображениями на сетчатке, а с участками внешнего зрительного пространства, т.е. рецептивные поля, в которых отражается внешнее зрительное пространство.
К этим нейронам поступает информация от верхнего двухолмия (о движениях глаз) и от зрительной коры (от …).
Эта область обеспечивает все ориентационные представления субъекта, т.е. расположение объектов вокруг субъекта и расположение самого субъекта относительно этих объектов.
Эта часть заднетеменной коры называется система «Где». С помощью нейронов этой области определяется расположение субъекта и определение его предметного окружения. Эта система имеет мощные анатомические связи со зрительной корой и с верхним двухолмием. Она посылает свои эфференты в префронтальную кору. Слияние внешней информации с переработанной информацией, которая даёт зрительной коре представление об относит.форм.предметов, обеспечивает константность изображения и зрительного поля.
Нижневисочная кора имеет такие же мощные анатомические связи со зрительной корой. В этой области находятся нейроны, которые обеспечивают категоризацию объектов, т.е. разделение на определённые классы, и соответственно, опознание объектов. Эта система называется система «Что», т.е. эти нейронные обеспечивают целостность восприятия.
Эфференты системы «Что» также направляются в префронтальную кору.
Поскольку гиппокамп связан с фиксацией следов памяти, в том числе и зрительных образов, то и категоризация связана с эталонами памяти. Функция этих областей связана со зрением.
И константность, и целостность зрительного восприятия формируется в ассоциативных областях: заднетеменной и нижневисочной.
ФАЙЛ 8
Свойства корковый нейронов
Это свойство пластичности присуще первичной зрительной коре, но это свойство ограничено чувствительным или сенситивным периодом у млекопитающих.
Детекторы формируются прижизненно в зависимости от воздействия окружающей среды. У котят – около 6 недель, у детей – 18 месяцев. В этот период формируются нейроны первичной зрительной коры ( …). Распознавание в зрительной коре характеристике образа называют подобразом (гештальтом), на уровне зрительной коры не обнаружено одного из важных свойств зрения – константности. В 1950-х гг. большинство учёных считали, что большинство органов чувств связано с ограниченным участками сенсорной коры, а вся остальная кора обеспечивает так называемые ассоциации. Накапливающие позднее экспериментальные данные заставили изменить эту точку зрения, поскольку для зрительной сенсорной системы кора, связанная с анализом информации, не ограничивается первичной зрительной корой. У млекопитающих с развитым зрением и приматов с переработкой зрительной информации связаны большие участки затылочной и височной долей. У макаков идентифицировано от 24 до 30 отдельных экстрастриарных участков зрительной коры. Многие из этих участков содержат карты поля зрения, связанные с различными характеристиками зрительных сигналов. Основной зрительный путь поступает в зрительную кору от сетчатки через коленчатое тело. Другой путь идёт в верхние бугры четверохолмия. В верхних буграх четверохолмия зрительная карта связана с контролем движения глаз и направлением взгляда. Эта структура состоит из 7 слоев клеток, они связаны не только со зрением. Сенсорный к верхнему двухолмию образует три карты: зрительную, соматосенсорную и слуховую, расположенные одна над другой. Зрительная карта формируется частично входом от сетчатки и частично из зрительной коры. Эта карта организована по другом принципу, чем в зрительной коре. Она представляет визуальное пространство вокруг животного. Другие сенсорные карты: соматосенсорная и слуховая – формируются по такому же принципу, что и зрительная, и располагаются точка в точку одна под другой.
…). Распознавание в зрительной коре характеристике образа называют подобразом (гештальтом), на уровне зрительной коры не обнаружено одного из важных свойств зрения – константности. В 1950-х гг. большинство учёных считали, что большинство органов чувств связано с ограниченным участками сенсорной коры, а вся остальная кора обеспечивает так называемые ассоциации. Накапливающие позднее экспериментальные данные заставили изменить эту точку зрения, поскольку для зрительной сенсорной системы кора, связанная с анализом информации, не ограничивается первичной зрительной корой. У млекопитающих с развитым зрением и приматов с переработкой зрительной информации связаны большие участки затылочной и височной долей. У макаков идентифицировано от 24 до 30 отдельных экстрастриарных участков зрительной коры. Многие из этих участков содержат карты поля зрения, связанные с различными характеристиками зрительных сигналов. Основной зрительный путь поступает в зрительную кору от сетчатки через коленчатое тело. Другой путь идёт в верхние бугры четверохолмия. В верхних буграх четверохолмия зрительная карта связана с контролем движения глаз и направлением взгляда. Эта структура состоит из 7 слоев клеток, они связаны не только со зрением. Сенсорный к верхнему двухолмию образует три карты: зрительную, соматосенсорную и слуховую, расположенные одна над другой. Зрительная карта формируется частично входом от сетчатки и частично из зрительной коры. Эта карта организована по другом принципу, чем в зрительной коре. Она представляет визуальное пространство вокруг животного. Другие сенсорные карты: соматосенсорная и слуховая – формируются по такому же принципу, что и зрительная, и располагаются точка в точку одна под другой.
В глубоких слоях четверохолмия образуется ещё одна карта – моторная. К ней из сенсорных карт поступает информация для контроля саккадических движений глаз, направляющих взгляд на важные части зрительного поля. В реализации этой функции принимает участие и фронтальная кора, т.е. лобное глазное поле. Волокна из верхних бугров четверохолмия через ядро подушки поступают в экстрастриарную зрительную кору. Подушка – это крупное ядро, лежит перед верхним бугром четверохолмия и образует заднюю часть таламуса. Это ядро направляет волокна в экстрастриарную кору и получает от неё афференты. Экстрастриарная кора получает афференты одновременно из стриарной коры и из подушки. Благодаря сложным связям зрительных областей обеспечивается целостная деятельность зрительной системы. При передаче информации от наружного коленчатого тела к стриарной коре крупноклеточные (М-типа) волокна несут сигналы о движении и форме, а парвоцеллюлярные (P-волокна) – о цвете и частично о форме. Причём эти волокна изолированы друг от друга и при входе в кору. Магноцеллюлярные волокна идут к клеткам, выделяющим края, а парвоцеллюлярные волокна – к клеткам с двойной цветовой оппонентностью и к клеткам, выделяющим положение контуров стационарных объектов.
Таким образом, в зрительной системе имеется три потока информации:
1. О форме, движении и стереоскопии;
2. О цвете;
3. Обо всех четырех характеристиках зрительного окружения.
Такие три раздельные потока информации поступаю и в экстрастриарную кору (поля 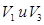 ). Это 18-е поле Бродмана. Зрительные путь прослежены до 19-го поля, содержащего области
). Это 18-е поле Бродмана. Зрительные путь прослежены до 19-го поля, содержащего области  и
и 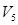 , и дальше во многие другие части коры, в том числе, теменной и височной. Таких участков в коре насчитывают до 30 и они связаны между собой волокнами в обоих направлениях. Информация из стриарной коры направляется в другие части мозга, в т.ч. в базальные ядра и в промежуточный мозг. Особенность: они связаны со зрительной корой и относятся к отдельным участкам. Наиболее важным для зрительной функции являются заднетеменная и нижневисочная. Эти зоны осуществляют сложную обработку зрительных сигналов, связанную с построением образа и переходом от описания отдельных признаков объектов к описанию предметов и отношений между ними. Восприятие сложных объектов, описание их частей и соотношения между ними проводится разными механизмами мозга. По данным современных исследований, эти механизмы локализованы в разных областях коры. Механизм классификации объектов и их инвариантное описание – в нижневисочной коре, а механизм описания пространственных отношений и полное описание образа – в заднетеменной коре. Эксперименты для подтверждения этих механизмов и их локализации были проведены на 3 группах обезьян. Первая группа контрольная, у второй группы билатерально удаляли нижневисочную кору, а у животных билатеральных удаляли заднетеменную кору. До проведения операции у всех животных вырабатывали пищевой двигательный условный рефлекс на дифференцировку зрительно предъявляемой фигуры, т.е. одну из 6 фигур, предъявляемых одновременно, подкрепляли. Эту процедуру проводили и после экстирпации коры. После упрочения рефлекса проводили преобразования фигур: меняли их размер и ориентацию.
, и дальше во многие другие части коры, в том числе, теменной и височной. Таких участков в коре насчитывают до 30 и они связаны между собой волокнами в обоих направлениях. Информация из стриарной коры направляется в другие части мозга, в т.ч. в базальные ядра и в промежуточный мозг. Особенность: они связаны со зрительной корой и относятся к отдельным участкам. Наиболее важным для зрительной функции являются заднетеменная и нижневисочная. Эти зоны осуществляют сложную обработку зрительных сигналов, связанную с построением образа и переходом от описания отдельных признаков объектов к описанию предметов и отношений между ними. Восприятие сложных объектов, описание их частей и соотношения между ними проводится разными механизмами мозга. По данным современных исследований, эти механизмы локализованы в разных областях коры. Механизм классификации объектов и их инвариантное описание – в нижневисочной коре, а механизм описания пространственных отношений и полное описание образа – в заднетеменной коре. Эксперименты для подтверждения этих механизмов и их локализации были проведены на 3 группах обезьян. Первая группа контрольная, у второй группы билатерально удаляли нижневисочную кору, а у животных билатеральных удаляли заднетеменную кору. До проведения операции у всех животных вырабатывали пищевой двигательный условный рефлекс на дифференцировку зрительно предъявляемой фигуры, т.е. одну из 6 фигур, предъявляемых одновременно, подкрепляли. Эту процедуру проводили и после экстирпации коры. После упрочения рефлекса проводили преобразования фигур: меняли их размер и ориентацию.
Группа животных с удалённой нижневисочной корой при изменении размера фигуры на 25% резко снижало вероятность правильного опознания, а если размер фигуры уменьшался в 2 раза, то выбор становился случайным. Такие же результаты получили в этой группе при изменении ориентации фигур.
Животные с удалённой заднетеменной корой опознавали предмет при изменении размера и ориентации, причём опознавали даже лучше, чем контрольная группа. Т.е. разрушение задней теменной коры не нарушало оценки, которые инвариантно описывали объект. Нарушалось только восприятие пространственных отношений. Разрушение нижневисочной коры нарушало описание зрительного образа, и в этом случае исчезала целостность восприятия. Оказалось, что такой механизм присутствует только у высших животных, его нет у низших. Нейроны этой области не изменяют реакции на подкрепляемые и неподкрепляемые стимулы. В целом, исследование нейронов нижневисочной коры показало, что они могут быть разделены на три группы:
1. Первая группа реагирует на сенсорное значение стимулов и не соотносит его с моторными программами;
2. Вторая группа неоднородна, часть из этих нейронов реагирует на любое появление стимула в поле зрения, а другая часть – только при сопровождении стимула актом внимания;
3. Третья группа нейронов реагирует на сложные признаки образа, инвариантные преобразованиям изображения.
Этот механизм выделения признаков нижневисочной корой был назван системой «Что». Эта система служит для опознания и категоризации объектов. Она начинается от ганглиозных клеток сетчатки, волокна которых поступают в наружное коленчатое тело, затем в поле  зрительной коры, далее в поле
зрительной коры, далее в поле  , и затем в нижневисочную кору. Отдельные признаки объекта обрабатываются параллельно в различных зонах коры. Локальное поражение одной из частей коры нарушает восприятие одного признака объекта. Например, ориентацию и длину линий, сложные признаки предмета, цвет и восприятие. Большая часть нейронов задней теменной коры реагирует на движение объектов в поле зрения и управляет саккадами. Предполагают, что в теменной коре локализованы механизмы, которые служат для обработки сенсорной информации и одновременно для генерации действий. Поэтому заднетеменной коре приписывают свойства создания модели окружающего пространства, описание локализаций и перемещение в этом пространстве объектов по отношению к телу наблюдателя. Кроме этого, область заднетеменной коры определяет положение и перемещение собственного тела в пространстве. Таким образом, механизмы заднетеменной коры соединяют два потока информации: об окружающем пространстве и о собственном положении наблюдателя в этом пространстве. Эта система получила название система «Где». Она начинается от клеток сетчатки, волокна которых поступают в верхние двухолмия. Верхние слои клеток двухолмия реагируют на зрительные образы, а глубокие слои запускают саккадические движения глаз. Амплитуда и направление саккад обеспечивают попадание объекта в центр поля зрения. Во время движения глаз зрительные сигналы смещаются по сетчатке, но образ зрительного мира не изменяется, потому что рецептивные поля нейронов двухолмия представлены участками зрительного поля, а не участками сетчатки. Зрительный образ дополняется сигналами о движении глаз. Эта система формируется после рождения в результате обучения, что подтверждено экспериментами на обезьянах.
, и затем в нижневисочную кору. Отдельные признаки объекта обрабатываются параллельно в различных зонах коры. Локальное поражение одной из частей коры нарушает восприятие одного признака объекта. Например, ориентацию и длину линий, сложные признаки предмета, цвет и восприятие. Большая часть нейронов задней теменной коры реагирует на движение объектов в поле зрения и управляет саккадами. Предполагают, что в теменной коре локализованы механизмы, которые служат для обработки сенсорной информации и одновременно для генерации действий. Поэтому заднетеменной коре приписывают свойства создания модели окружающего пространства, описание локализаций и перемещение в этом пространстве объектов по отношению к телу наблюдателя. Кроме этого, область заднетеменной коры определяет положение и перемещение собственного тела в пространстве. Таким образом, механизмы заднетеменной коры соединяют два потока информации: об окружающем пространстве и о собственном положении наблюдателя в этом пространстве. Эта система получила название система «Где». Она начинается от клеток сетчатки, волокна которых поступают в верхние двухолмия. Верхние слои клеток двухолмия реагируют на зрительные образы, а глубокие слои запускают саккадические движения глаз. Амплитуда и направление саккад обеспечивают попадание объекта в центр поля зрения. Во время движения глаз зрительные сигналы смещаются по сетчатке, но образ зрительного мира не изменяется, потому что рецептивные поля нейронов двухолмия представлены участками зрительного поля, а не участками сетчатки. Зрительный образ дополняется сигналами о движении глаз. Эта система формируется после рождения в результате обучения, что подтверждено экспериментами на обезьянах.
FILE 9
Лекция 12
Звуковые волны представляют собой передаваемые от источника звука механические смещения молекул воздуха или другой упругой среды. Скорость распространения звуковых волн в воздухе - 343 м/с при 0° Цельсия. В воде и металлах эта скорость значительно выше. Закономерно чередующиеся участки сжатия и разряжения молекул среды описываются синусоидами, которые отличаются амплитудой и частотой. При суперпозиции (наложении) звуковых волн с различными амплитудами и частотами они наслаиваются друг на друга, образуя комплексные волны.
Физическим понятиям амплитуды и частоты соответствуют субъективные ощущения высоты и громкости. Звук, образованные синусоидальными колебаниями одной частоты, вызывает ощущение определённой высоты и обозначается как тон. Комплексные волны состоят из одного тона (это самые низкие частоты) и обертонов (или гармоник), представляющие более высокие частоты. В повседневной жизни тоны всегда бывают комплексными (они состоят из нескольких синусоид). Индивидуальное сочетание комплексных волн определяет характерный тембр человеческого голоса или музыкального инструмента. Слуховая система человека способна различать высоту звука только у периодических звуковых сигналов, а звуковые раздражители, состоящие из беспорядочного сочетания частотных и амплитудных компонентов, воспринимаются как шум. Дети воспринимают звуковые волны в диапазоне 16-20.000 Гц, но приблизительно с 15-20 лет диапазон частотного восприятия суживается, так как утрачивается чувствительность звуковой системы к самым высоким звукам. Ухудшение качества звукового восприятия у пожилых людей является естественным процессом. В норме, независимо от возраста, человек легче всего воспринимает звуковые волны в диапазоне 100-2000 Гц. Это особенно важно, поскольку речь и музыкальные инструменты звучат именно в этом диапазоне.
Нижняя нота рояля - 27,5 Гц - 82,4 м
Нижний певческий бас - 80 Гц - 2,7 м
Нота ЛЯ - 440 Гц - 79 * 10^(-2) м
Высокие тенор - 500 Гц - 69*10^(-2) м
Альт - 650 Гц - 78 * 10^(-2) м
Верхняя ДО - 1049 Гц - 33 * 10^(-2) м
Колоратурное сопрано 1350 Гц - 25*10^(-2) м
Самая высокая нота рояля - 4186 Гц - 8,2 * 10^(-2) м
Чувствительность слуховой системы к минимальным изменениям высоты звука определяется разностным порогом частоты. В оптимальном для восприятия диапазоне 1000 Гц - этот порог составляет примерно 3 Гц. При изменении высоты звука до 2000 Гц разностный порог повышается до 6 Гц, а при 3000 Гц разностный порог 9 Гц и т.д. Таким образом, получается логарифмическая шкала.
Амплитуда звуковых волн определяет величину звукового давления, под которой понимают силу сжатия, действующую на перпендикулярно расположенную к ней площадь. Акустическим эталоном, близким к абсолютному порогу слухового восприятия принято считать величину 2*10^(5) единиц давления / кв. метр. Сравнительной единицей изменения громкости считают Дб (децибел) и громкость измеряется по логарифмической шкале, где логарифм десятичный, он высчитывается как отношение действующего звукового давления к эталонному давлению. Эта величина умножается на 20.
В Дб также измеряют интенсивность различных источников звука. При этом под ней понимают мощность или плотность звуковых волн в единицу времени. За эталонную интенсивность принимают величину 10^(-12) Вт / кв. м.
Количество Дб для измеряемой интенсивности определяют по формуле:
| lg (измеряемая интенсивность / эталонная интенсивность) * k, k=10 |
Субъективно воспринимаемая громкость звучания зависит не только от уровня звукового давления, но и от частоты звукового стимула. Чувствительность слуховой системы максимальна для раздражителей с частотами 500-4000 Гц. При действии звуков менее 500 Гц или более 4000 Гц порог звуковой чувствительности повышается.
В наружном, среднем и внутреннем ухе происходят процессы, необходимые для слухового восприятия. Эти процессы подготовительные, они оптимизируют параметры передаваемых звуковых колебаний при одновременном сохранении характера сигналов.
Преобразование энергии в рецепторные потенциалы волосковых клеток. Ушные раковины состоят из хряща, покрытого кожей и соединенного с хрящом наружного слухового прохода. Внутренность слухового прохода также выстлана кожей. в ней содержатся сальные железы и волосяные мешочки.
Движения ушных раковин млекопитающих в направлении источника звука помогают обнаружить источник и его пространственное расположение. Как правило человек не может двигать ушными раковинами. Сохранение способности к движению ушными раковинами расценивается как атавизм.
Функцию определения расположения источника звука у человека выполняет рельеф ушных раковин и их расположение на противоположных сторонах головы. Такое положение ушных раковин позволяет различать поступление звука спереди и сзади. Наружный слуховой проход заканчивается барабанной перепонкой, представляющей собой вогнутую в полость среднего уха соединительно-тканную перегородку толщиной около 0,1 мм. Звуковые волны вызывают колебание барабанной перепонки вокруг оси, проходящей поблизости от оси проходящей вблизи его верхнего края. У барабанной перепонки нет собственного периода колебания. Её волокна способствуют колебания, повторяющие частоту действующих звуковых волн. С той же частотой барабанная перепонка передаёт колебания присоединённым к ней слуховым косточкам.
FILE 10
Чувство равновесия
Органы вестибулярной системы:
- Сферический мешочек (саккулюс)
- Эллиптический мешочек (утрикулюс)
- Полукружные каналы
Лабиринт некоторых позвоночных:
- Утрикулус
- Саккулус
- Лагена
- Базилярный сосочек
- Базилярная мембрана
- Улитка
- Первоначально ухо являлось только органом равновесия.
- Лишь на поздних стадиях эволюции оно стало органом слуха.
- Улитка — это вырост сферического мешочка.
Сферический и эллиптический мешочки
- Сферический и эллиптический мешочки заполнены жидкостью — эндолимфой.
- На внутренней поверхности мешочка находятся пятна, называемые макулами. Макула эллиптического мешочка — лежит горизонтально, а макула сферического — вертикально.
- Каждая макула содержит опорные и волосковые клетки. Волосковые клетки являются рецепторами вестибулярной системы.
- Поверхность макулы покрыта желеобразным веществом, сверху которого находятся отолиты.
Отолиты («слуховые камешки», отокинии, статолиты) — плотные кристаллы карбоната кальция.
Размер — 0,5-10 мкм.
Когда мы наклоняем голову, отолиты под действием гравитации скатываются вниз и тянут за собой слой желеобразного вещества.
Закидывание головы назад. Пороги различения наклона головы:
- В сторону — около 1 градуса
- Вперёд/назад — около 2 градусов.
Чувство гравитации
Эллиптический мешочек. Детектирует движение тела по горизонтали (вперёд/назад).
Сферический мешочек. Детектирует движение тела по вертикали (вверх/вниз).
Вестибулярные мешочки стимулируются силой тяжести.
Благодаря им у нас появляется чувство гравитации — мы ощущаем где находится верх, а где низ.
Полукружные каналы — заполненные эндолимфой дугообразные трубки, расположенные в трёх взаимно перпендикулярных плоскостях.
Различают:
- передний канал;
- задний канал;
- латеральный канал.
Каждый канал заканчивается расширением и виде сферической камеры — ампулы.
При вращении головы (тела) жидкость в полукружных каналах смещается и наклоняет купол в ампуле.
Порог чувствительности — 2-3 градуса / кв. сек
Принцип работы купола (рисунок)
Чувство вращения
Куполы в ампулах полукружных каналов детектируют только начало и окончание вращательного движения.
В норме на силу тяжести куполы не реагируют.
Содержащийся в алкоголе этанол изменяет плотность эндолимфы полукружных каналов. В результате купол начинает также реагировать на силу тяжести. Поэтому в результате сильного опьянения возникает ощущение головокружения.
Вестибулярный нерв
Афферентные волокна от волосковых клеток вестибулярных мешочков и полукружных каналов направляются в ганглий Скарпы (вестибулярный ганглий), где находятся тела нейронов. Аксоны этих нейронов (18-20 тыс. волокон) образуют вестибулярный нерв (VIII), который направляется в вестибулярный комплекс продолговатого мозга.
Ганглий Скарпы находится рядом со слуховым проходом и его можно стимулировать.

