




Між організмами в екосистемах існують трофічні зв'язки, на основі яких формуються ланцюги живлення. Першою ланкою будь-якого ланцюга живлення є рослини, які утворюють органічну речовину в процесі фотосинтезу. Рослинами живляться рослиноїдні тварини, останніми - дрібні хижаки, які, у свою чергу, стають здобиччю більших за розміром хижаків. Характерною ознакою цього ланцюгу живлення є те, що із зростанням його рівня збільшується розмір тварин і зменшується їх кількість, наприклад, трава-слимак-жаба-лисиця. Такі ланцюги живлення називають ланцюгами виїдання. У трофічних ланцюгах паразитів, навпаки, зміни відбуваються від більших за розміром і менш чисельних організмів до менших за розмірами і більш чисельних, наприклад, тварина-блохи-бактерії-віруси. Приклади ланцюгів живлення наведені в табл.3.1, з якої видно різницю між звичайним ланцюгом виїдання та ланцюгом паразитів.
В екосистемах може бути кілька паралельних ланцюгів живлення, між якими існують зв'язки, адже організми живляться різними об'єктами і самі є поживою для різних членів екосистем. В реальних умовах ланцюги живлення можуть перехрещуватись і утворювати мережу живлення. Саме складна система трофічних ланцюгів забезпечує цілісність і динамічність екосистем. Різні рівні живлення в екологічній системі називаються трофічними.
Трофічний рівень - це сукупність організмів, об'єднаних типом харчування. До першого трофічного рівня належать автотрофні організми; на другому рівні - рослиноїдні тварини (консументи І порядку); на третьому - тварини, які харчуються рослиноїдними тваринами (консументи II порядку, первинні хижаки); на четвертому рівні - вторинні хижаки (консументи III порядку). Замикають трофічний ланцюг редуценти, які можуть займати всі рівні, починаючи з другого.
Таблиця 3.1
Природні ланцюги живлення (трофічні рівні)
| Продуценти | Консументи | ||
| Рослиноїдні (І порядку) | Первинні хижаки (II порядку) | Вторинні хижаки (III порядку) | |
| Рослини | Лось, олень, косуля Заєць | Ведмідь, вовк, лисиця Лисиця, борсук, тхір, горностай, канюк | Блохи, іксодові кліщі |
| Слизун, слимак | Жаба, ящірка, гадюка, їжак | Лисиця, канюк |
Трофічні ланцюги ґрунтуються на другому законі термодинаміки, згідно з яким певна частина енергії завжди втрачається і стає недосяжною для використання (зростання ентропії). У відповідності із законом піраміди енергії (Р. Ліндеман) з одного трофічного рівня екологічної піраміди на інший переходить у середньому не більше 10% енергії, а зворотний потік енергії (з більш високих рівнів на нижчі рівні) набагато слабший (не більше 0,5-0,25%). При цьому, кількість консументів трофічного рівня обмежується правилом екологічної піраміди.
Продуктивність екологічних систем і співвідношення в них різних трофічних рівнів зображують, як правило, у вигляді пірамід. Сьогодні, розрізняють три основні типи екологічних пірамід:
1) піраміди чисел, які відображають чисельність окремих організмів ланок трофічного ланцюга;
2) піраміди біомаси, які характеризують загальну суху масу ланок трофічного ланцюга;
3) піраміди енергії, які показують величину потоку енергії або
продуктивність кожного трофічного рівня в ланцюгу живлення.
Це чітко показано на прикладі екологічної піраміди Ю. Одума (рис. 3.2). Вихідною продукцією трофічного ланцюгу є посіви люцерни на площі 4 га. Цього достатньо для годівлі 4-5 телят (припускається, що вони вживають тільки люцерну). Телятину споживає 12-річний хлопчик (припускається, що він вживає тільки телятину). Люцерна використовує 0,24% сонячної енергії, що надійшла на її посіви, телята - 8% енергії люцерни, хлопчик - лише 0,7% енергії, накопиченої телятами. Отже, для існування хлопчика протягом року достатньо близько 1/1000000 частини сонячної енергії, що надійшла на 1 га посіву люцерни.
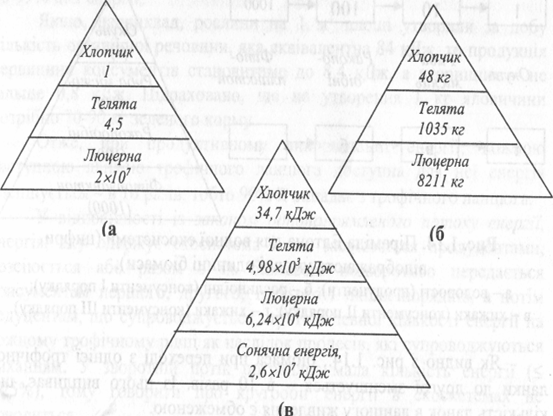
Рис. 3.2. Екологічна піраміда Ю. Одума: а - піраміда чисел; б - піраміда біомас; в - піраміда енергій.
Ю.Одум (1986) на підставі аналізу цих пірамід сформулював наступне екологічне правило: дані піраміди чисел призводять до перебільшення ролі малих організмів, а дані піраміди біомас - великих. У малих організмів більш інтенсивний метаболізм, тому лише потік енергії є найбільш придатним показником для порівняння будь-якого компоненту з іншим і всіх компонентів між собою.
Аналіз піраміди енергій дозволяє не тільки оцінити ефективність використання енергії екосистемою, але і порівнювати різні екосистеми, і визначати відносну роль трофічної ланки. Знання законів продуктивності екологічних систем має надзвичайно велике практичне значення, зокрема для сільського господарства. Це дає можливість розраховувати необхідну площу орних земель, визначати стратегію розвитку сільського господарства. Наприклад, найбільш дешевою є продукція рослинництва; утримання хижих тварин є дорогим, тощо.
Одним із завдань загальної екології є визначення меж стійкості екосистем до зовнішніх впливів, в тому числі антропогенного. Під стійкістю екосистем розуміють здатність її до реакцій, які пропорційні по величині силі впливу, але напрямлені протилежно. При перевищенні певної критичної величини впливу екосистема втрачає стійкість.
На стійкість екологічних систем до зовнішніх дій впливають ряд факторів, зокрема ефективність використання енергії в екосистемі, біорізноманіття, тощо. Основою стійкості екосистем є біорізноманіття. Існують певні закономірності відносно стійкості системи до зовнішніх впливів:
- чим більша розмаїтість біоценозу, тим більша екологічна стійкість екосистеми;
- чим більш різноманітними умовами характеризуються межі екосистем, тим більше видів містить відповідний біоценоз, тощо.
Так, у відповідності із принципом Ле Шательє-Брауна, при зовнішній дії на екологічну систему, що виводить її із стану рівноваги, ця рівновага зміщується в напрямку послаблення ефекту зовнішньої дії. Отже, екосистеми є динамічними. Пом'якшення зовнішнього впливу на екосистему може відбуватись за рахунок різноманітних процесів, зокрема, це зміна кругообігу речовин, зміна видо-спеціалізації організмів та їх функціональній ролі в системі, тощо.
Ю.А.Ізраель для оцінки стійкості екосистем і біосфери щодо природних явищ і антропогенних порушень запропонував поняття про екологічний резерв екосистеми, під яким розуміють різницю між граничне допустимим відхиленням та фактичним станом екосистеми, тобто це так званий запас міцності (буферність).
У відповідності із законом максимальної енергії та інформації (Ю. Одум, Г. Одум, М. Реймерс) у конкуренції з іншими екосистемами зберігається та, яка найбільше сприяє надходженню енергії та інформації і використовує їх кількість найефективніше.
Для нормального функціонування, екосистеми утворюють накопичувачі високоякісної енергії, яка витрачається на забезпечення надходження нової енергії, забезпечення кругообігу речовини створення механізмів регулювання І підтримки стійкості системи, її здатність пристосовуватись до змін. Саме такі екосистеми є стійкими.
Стійкість екосистем визначається її асимілюючою ємністю, під якою розуміють показник максимальної кількості забруднюючої речовини, яка може бути за одиницю часу накопичена, зруйнована, трансформована і виведена за межі екосистеми без порушення нормального її функціонування. Ця величина залежить від багатьох факторів, як природних, так і антропогенних. Наприклад, за рахунок значного виносу речовин за межі екосистеми річок, їх асимілююча ємність є високою, тощо. Чим більшою є екосистеми, тим більшою асимілюючою ємністю вона характеризується. В той же час, не слід забувати, що екосистеми є цілісними системами і порушення їх цілісності призводить до порушення механізмів саморегуляції. Згідно закону екологічної кореляції, в екосистемі всі живі компоненти та абіотичні екологічні компоненти функціонально відповідають одне одному і випадання однієї частини системи неминуче викличе випадання інших частин екосистеми і призведе до функціональних змін. Жодна частина екосистеми не може існувати без іншої.
Отже, екосистеми є стійкими динамічними системами з власними механізмами саморегуляцій. Саме на цьому ґрунтується природна екологічна рівновага і тому екосистеми можуть існувати тривалий час.

