




МЕТОДИЧЕСКИЕ УКАЗАНИЯ
ДЛЯ САМОСТОЯТЕЛЬНОЙ РАБОТЫ СТУДЕНТОВ ПРИ ПОДГОТОВКЕ К ПРАКТИЧЕСКОМУ ЗАНЯТИЮ
| Учебная дисциплина | “ФИЗИОЛОГИЯ” |
| Модуль № 1 | “ОБЩАЯ ФИЗИОЛОГИЯ” |
| Содержательный модуль №2 | Возбудимые структуры. |
| Тема практического занятия №3 | Исследование проведения возбуждения нервными волокнами и через нервно-мышечный синапс. Исследование потенциала действия целостных нервов и мышц |
| Курс | 2-й |
| Факультет | Медицинский |
Киев – 2006
1.Актуальность темы:
Механизмы проведения возбуждения нервными и мышечными волокнами и через нервно-мышечный синапс – это способ передачи информации. Эти закономерности имеют не только теоретический интерес. Врачу часто приходится наблюдать нарушение проведения возбуждения нервными и мышечными волокнами и через нервно-мышечные синапсы и целенаправленно влиять на эти процессы с помощью фармакологических средств.
2.Учебные цели:
Ø Объяснять механизмы проведения возбуждения нервными и мышечными волокнами.
Ø Анализировать роль основных факторов, которые определяют скорость проведения возбуждения нервными и мышечными волокнами.
Ø Трактовать закономерности проведения возбуждения нервными и мышечными волокнами.
Ø Объяснять и анализировать механизм формирования и свойства ПД, которые отводятся от целостных нервов и мышц; механизмы формирования электронейрограммы и электромиограммы.
Ø Объяснять и анализировать механизмы и закономерности проведения возбуждения через нервно-мышечные синапсы и возможности блокады нервно-мышечной передачи.
Базовый уровень подготовки
Студенты имеют навыки из предыдущих учебных дисциплин:
| Названия предыдущих дисциплин | Получены навыки |
| Анатомия человека | Анализировать информацию о строении тела человека, систем, что его составляют, органов и тканей |
| Гистология, цитология, эмбриология | Интерпретировать микроскопическую и субмикроскопическую структуру клеток |
| Медицинская и биологическая физика | Трактовать общие физические и биофизические закономерности, которые лежат в основе жизнедеятельности человека |
| Медицинская биология | Объяснять закономерности проявлений жизнедеятельности человеческого организма на молекулярно-биологическом и клеточном уровнях. |
Задание для самостоятельной работы во время подготовки к практическому занятию
Перечень основных терминов, параметров, характеристик, которые должен усвоить студент при подготовке к практическому занятию
| Срок | Определение |
| Нервно-мышечный синапс | Место контакта нервного окончания двигательного нерву с мышечным волокном |
| Медиатор (нейромедиатор) | Химическое вещество, которое выделяется из нервного окончания нейрона через его пресинаптическую мембрану и осуществляет передачу информации через синаптическую щель к следующей структуре синапса – постсинаптической мембране. |
| Н-холинорецепторы | Поверхностные белки постсинаптической мембраны, с которыми взаимодействует медиатор ацетилхолин |
| Потенциал концевой пластинки (ПКП) | Местный деполяризационный потенциал, который возникает на постсинаптической мембране (концевой пластинке) нервно-мышечного синапса при взаимодействии медиатора ацетилхолина с Н-холинорецепторами концевой пластинки (КП), благодаря этому увеличивается проницаемость КП для ионов натрия через хемозависимые натриевые каналы. |
Теоретические вопросы
1) Механизмы проведения возбуждения (ПД) нервными и мышечными волокнами, особенности проведения возбуждения миелиновыми нервными волокнами.
2) Факторы, которые определяют скорость проведения ПД нервными и мышечными волокнами. Фактор надежности. Классификация нервных волокон в зависимости от их диаметра.
3) 3акономерности проведения возбуждения нервными и мышечными волокнами, их значение для передачи информации.
4) ПД целостных нервов и мышц. Механизм их формирования и свойства. Общие представления об отведении ПД от объемного проводника.
5) Электромиография, механизмы формирования электромиограммы.
6) Механизмы и закономерности проведения возбуждения через нервно-мышечный синапс. Медиатор, мембранные циторецепторы и блокаторы нервно-мышечных синапсов.
Практические работы
1) Исследование свойств суммарных ПД изолированного нерва.
2) Исследование скорости проведения возбуждения изолированным нервом.
3) Исследование механизмов проведения возбуждения через нервно-мышечные синапсы.
Содержание темы
1) Механизмы и закономерности проведения ПД по нервным и мышечным волокнам представлено на схеме.
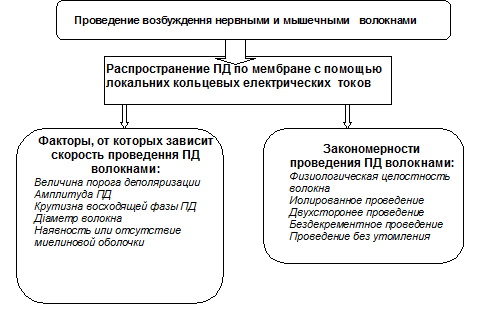
ПД, который генерируется в определенном участке нервного волокна, действует как стимул на соседний участок мембраны нервного волокна благодаря местным кольцевым электрическим токам, которые возникают между этими участками мембраны нервного волокна.
Скорость распространения ПД по нервному волокну зависит от типа нервного волокна. Она больше в миелиновых нервных волокнах, потому что миелиновая оболочка является изолятором для электрического тока и потому ПД возникает только в перехватах Ранвъе - это сальтаторный (скачкообразный) вид проведения ПД. Она меньше в немиелинизированных нервных волокнах, потому что благодаря местным электрическим токам ПД возникает в соседнем участке мембраны на малом расстоянии - это непрерывный вид проведения импульса. Так же проводится возбуждение по мембране мышечного волокна.
Скорость проведения пропорциональна диаметру нервного волокна. В миелиновых нервных волокнах диаметр составляет от 1 до 20 мкм, а скорость проведения нервного импульса: 6-120м/с. В немиелиновых нервных волокнах диаметр составляет около 1 мкм, а нервный импульс распространяется со скоростью от 0,5 м/с до 2,5м/с.
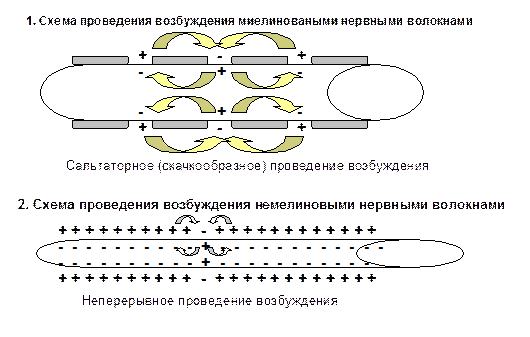
Параметры импульса постоянного электрического тока, которые будут вызывать генерацию ПД:
На поверхности мембраны должен располагаться катод (-), а в клетке - анод (+) - это "катодное направление электрического тока", потому что только при уменьшении МПС открываются потенциало-зависимые активационные ворота (m) натриевых каналов:

Величина электрического тока, который действует на мембрану, должна быть не меньше пороговой величины (ΔЕ), потому что только в таком случае открываются потенциало-зависимые активационные ворота (m) всех натриевых каналов, которые имеет мембрана, и возникает ПД.
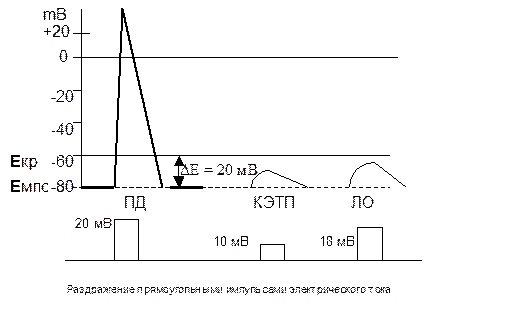
- При действии на нервное волокно допорогового электрического тока ПД не возникает, а возникают местные потенциалы:
- Катэлектротонический потенциал (КЭТП) возникает благодаря пассивному действию электрического тока катодного направления на мембрану нервного волокна.
- Локальный ответ (ЛО) - обусловленный входом ионов натрия через некоторые натриевые каналы при их активации, но большинство потенциало-зависимых ворот натриевых каналов закрыто при допороговой величине электрического тока.
- Местные потенциалы не распространяются на значительное расстояние.
- Они подчинены "закону силовых отношений": чем больше сила допорогового электрического тока, тем больше величина местного потенциала.
Длительность импульса электрического тока должна быть не меньше пороговой величины (ΔЕ), что связано с характеристикой мембраны - электрическая модель мембраны имеет емкостное (С) и омическое (R) сопротивление, а время действия электрического тока, на эту систему должен быть не меньше постоянного времени мембраны (t):
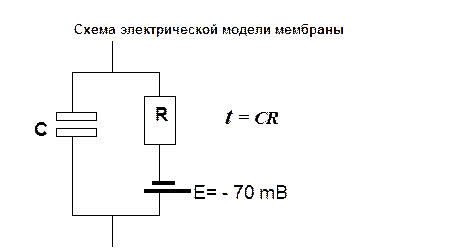
Скорость (крутизна) развития импульса постоянного электрического тока должна быть не меньше пороговой величины (1), потому что при медленной скорости нарастания электрического импульса (2) происходит инактивация натриевых каналов (закрытие потенциалозависимых h-ворот) натриевых каналов, что приводит к повышению уровня Екр (процессу аккомодации) и ПД не возникает.
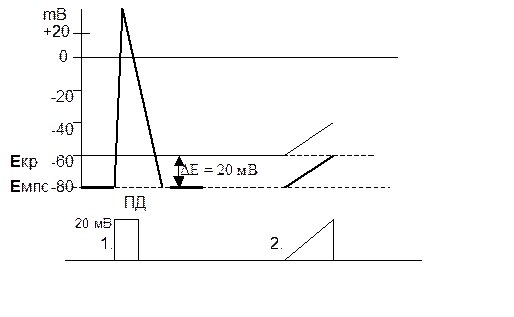
Физиологические механизмы местной и проводниковой анестезии.
Местные и проводниковые анестетики (прокаин, тетракаин) действуют непосредственно на активационные (m-ворота) натриевых каналов, затрудняя их открывание и, тем самым, уменьшая возбудимость.
Когда возбудимость уменьшается настолько, что соотношение между величиной ПД и пороговым потенциалом ("фактор надежности") становится меньше 1, нервный импульс не проводится через анестезированный участок мембраны.
Анестетики связываются только с открытыми каналами в участке между входом канал и "m"-воротами. Закрытие натриевых каналов длится несколько миллисекунд, но повторяется с высокой частотой, потому вход ионов натрия в клетку становится неэффективным.
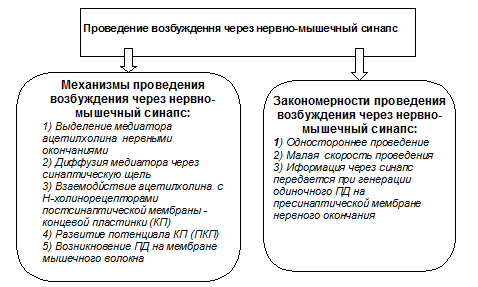
2) Механизмы и закономерности проведения возбуждения через нервно-мышечный синапс поданы на схеме:
Нервно-мышечный синапс – это место контакта аксона нервного окончания двигательного нерва и мышечного волокна, через который передается информация от аксона к мышечному волокну.

Медиатор ацетилхолин, который обеспечивает передачу информации в нервно-мышечном синапсе; взаимодействует с Н-холинорецепторами КП.
Синтез и сохранение медиатора ацетилхолина происходит в нервных окончаниях аксона из холина и ацетилкоензима А при участии фермента ацетилтрансферазы. Синтезированнный ацетилхолин сохраняется вместе с АТФ и протеогликаном в синаптических пузырьках: 5000 - 10000 молекул ацетилхолина в 1 пузырьке.
Проведение возбуждения через нервно-мышечный синапс имеет такие шаги:
Деполяризация пресинаптической мембраны возникает благодаря ПД, который распространяется по мембране аксона к нервному окончанию. Деполяризация приводит к открыванию ворот Са2+- каналов в пресинаптической мембране, через которые проходят ионы кальция в нервное окончание путем диффузии.
Повышение концентрации Са2+ в нервном окончании приводит к выходу медиатора (экзоцитозу) в синаптическую щель.
Ацетилхолин путем диффузии доходит до КП, где взаимодействует с Н-холинорецепторами, которые расположены в КП, в результате чего открываются хемозависимые ворота натриевых и калиевых каналов КП, что приводит к ее деполяризации, потому что ионов натрия через ионные каналы входит в клетку больше, чем выходит из клетки ионов калия благодаря большему электрохимическому градиенту для ионов натрия.
Деполяризация концевой пластинки имеет название “потенциал концевой пластинки” (ПКП), его величина достигает около 50 мВ. Это местный потенциал, который вызывает местные электрические токи между КП и мембраной мышечного волокна рядом с синапсом. Благодаря местным электрическим токам на мембране мышечного волокна генерируется ПД, распространение которого вызывает функцию мышечного волокна - его сокращение. Величина ПКП в 3 раза больше, чем пороговый потенциал мембраны мышечного волокна.
Содержание ацтилхолина в одной везикуле (пузырьке) достаточно, чтобы вызывать миниатюрные потенциалы КП (МПКП), но их величина допороговая и они не могут вызывать генерацию ПД в мышечном волокне. МПКП возникают спонтанно и возможно имеют трофическое влияние на мышечное волокно.
Деградация ацетилхолина.
Медиатор ацетихолин распадается на КП до ацетилкоензима А и холина под воздействием фермента ацетилхолинэстеразы (АХЭ). Половина холина, который образовался, возвращается через пресинаптическую мембрану в нервное окончание аксона вторичным активным транспортом, совмещенным с транспортом ионов натрия.
Ингибиторы АХЭ (неостигмин, пиридостигмин, амбеноний) блокируют деградацию ацетилхолина, продлевая длительность его действия и увеличивая величину ПКП.
Гемихолин блокирует транспорт холина через пресинаптическую мембрану, уменьшая его запасы в нервном окончании.
Физиологические механизмы миастении гравис (myastenia gravis).
Это заболевание характеризуется слабостью мышц и является следствием уменьшения количества Н-холинорецепторов в КП, что вызвано влиянием антител к этим рецепторам.
Применение ингибиторов АХЭ продлевает действие ацетилхолина на КП и частично компенсирует уменьшение количества рецепторов.

