




Изучение функциональной структуры фитоценозов направлено на установление взаимосвязей его компонентов, на.выяснение механизмов его функционирования. К сожалению, данный аспект структуры фитоценозов пока разработан еще очень слабо. В.В. Мазинг (1969) намечает следующие основные направления анализа функциональной структуры фитоценозов: 1)'изучение процессов переноса вещества, энергии и информации; 2) системно-структурный анализ фитоценозов, 3) изучение изменения функциональной структуры в пространстве и во времени, 4) качественная и количественная характеристика всех связей и процессов в фитоценозах, 5) индуктивный и дедуктивный анализ функциональной структуры.
В качестве элементов функциональной структуры фитоценоза нередко рассматривают элементы других аспектов структуры: ценопопуляции, синузии, микроценозы, конгрегации, микрогруппировки и т.д. Каждый из этих ценоэлементов характеризуется своей системой отношений между растениями, своей связью с местообитанием, особым обменом веществ и энергии, занимает особую экологическую нишу. Поэтому они выступают как более или менее обособленные функциональные блоки фитоценоза, как его подсистемы и, будучи связанными друг с другом образуют единую фитоценотическую систему. Однако существуют и чисто функциональные элементы структуры фитоценоза, к которым относят ценоячейку и копсорцию.
Под ценоячейкой B.C. Ипатов (1966) предложил понимать группу растений, функционально связанных друг с другом через среду Растения взаимодействуют в ценоячсйкс через их фитогенные поля- поэтому центрами ценоячеек выступают в первую очередь растения-эдификаторы, создающие вокруг себя достаточно выраженные фитогенные поля.
По мнению А.А. Ниценко (1971), в ценоячейке практически каждое растение влияет на каждое по схеме:

Вместе с тем ценоячейки не имеют фиксированных границ. Объясняется это тем, что одни и те же растения могут входить в состав разных соседних ценоячеек, Более того около каждого растения в фитоценозе существует своя ценоячейка и поэтому объем ценоячейки будет зависеть от того, какое растение мы выберем в качестве центрального. Следовательно, соседние ценоячейки частично перекрывают друг друга, в результате в фитоценозе образуется непрерывная ткань взаимовлияний (Ниценко, 1971), и это служит одним из источников возникновения непрерывности растительного покрова (Миркин, Розенберг, 1983).
Однако такая ситуация может наблюдаться лишь в фитоценозах, образованных видами одной жизненной формы, например, на лугах, где все растения обладают сопоставимыми по объему фитогенными полями, которые у соседних особей в значительной степени перекрывают друг друга. Иная картина наблюдается в фитоценозах, образованных видами разных жизненных форм, например, в лесах, где центрами ценоячеек являются отдельные деревья, имеющие достаточно обширные фитогенные поля. В данном случае ценоячейку образует дерево со всеми растениями подлеска и напочвенного покрова, находящимися в пределах фитогенного поля дерева (рис. 23). Очевидно, что в такой ценоячейке будут взаимодействовать друг с другом не все растения подчиненных ярусов, а только соседние растения, фитогенные поля которых в той или иной степени перекрываются.
Таким образом, объем ценоячейки определяется объемом фитогенного поля одного из взаимодействующих, или нескольких близко расположенных растений, занимающих в ней господствующее положение. Чем больше роль главного центрального растения, тем сильнее зависят от него другие растения и тем ценоячейка будет целостнее.
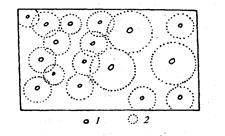
Рис. 23. Схема расчленения участка лесного фитоценоза на ценоячейки (Корчагин, 1976): 1 - стволы деревьев, 2 - кроны деревьев.
Консорция является элементом функциональной структуры биоценоза, включающего фитоценоз в качестве основного компонента. Различают два аспекта в понимании консорции - понятие консорции на индивидуальном уровне (на уровне особи) и на ценопопуляционном уровне.
Под индивидуальной консорцией, представление о которой было сформулировано В.Н. Беклемишевым (1951), понимают группу организмов в биоценозе, включающую автотрофное растение и все связанные с ним трофическими или прямыми топическими отношениями особи растений и животных. Примером такой консорции является дерево сосны вместе с ее микоризными грибами на корнях, с эпифитными мхами и лишайниками на стволе и ветвях, с паразитическими грибами в различных тканях, со всем множеством населяющих ее членистоногих.
Под ценопопуляционной консорцией Л.Г. Раменский (1952) и JE.M. Лавренко (1959) предложили понимать совокупность ценопопуляций вида высшего растения (автотрофа) в любом конкретном фитоценозе и связанных с ней трофически или прямыми топическими отношениями популяций растений и  животных. Примером такой консорции является совокупность ценопопуляций ели в еловом лесу и связанных с ней популяций паразитов, сапрофитов, лиан, эпифитов, симбионтов, вредителей, переносчиков пыльцы и семян и т.д.
животных. Примером такой консорции является совокупность ценопопуляций ели в еловом лесу и связанных с ней популяций паразитов, сапрофитов, лиан, эпифитов, симбионтов, вредителей, переносчиков пыльцы и семян и т.д.
Важнейшим свойством консорции как функциональной системы является наличие непосредственных (прямых) связей ее компонентов с центральным растением, Основное значение имеют такие типы трофических связей, как паразитический, сапрофитный, симбиотический, разнообразные трофические связи автотрофных растений с насекомыми-фитофагами, а также эпифитные связи.
Каждая консорция состоит из двух частей: центрального растения, или центральной ценопопуляции - ядро консорции и связанных с ним организмов, или популяций растений и животных. Компоненты консорции называют консортами, а центральный компонент - консортом-детерминантом, В качестве консортов-детерминантов могут выступать, по-видимому, все автотрофные растения, среди которых основную роль играют высшие растения, а среди последних важнейшими являются доминанты и эдификаторы.
Консорты-детерминанты являются продуцентами, они производят основную часть органическою вещества в биоценозе. Остальные консорты представлены в основном гетероторофами и в процессе своей жизнедеятельности потребляют органическое вещество, созданное продуцентами. Гетеротрофные консорты делятся на две группы - консументы и редуценты. К консументам, или биофитотрофам относятся организмы, потребляющие вещество и энергию живых автотрофных растений, а к редуцентам, или сапрофитотрофам - организмы, потребляющие отмершие части растений (детрит), минерализующие мертвое органическое вещество в процессе своей жизнедеятельности и использующие заключенную в нем энергию. Третья группа консортов представлена эпифитами и лианами, которые, являясь автотрофами, сами синтезируют органическое вещество, а консорт-детерминант используют как место для своего прикрепления или как опору.
Н.В. Беклемишев (1951) и Л.Г. Раменский (1952) под консорцией понимали совокупность организмов, или популяций, объединенных прямыми, непосредственными связями. Однако некоторые ботаники (Мазинг, 1966; Работнов, 1970 и др.) расширяют объем консорции, включая в нее также организмы (или популяции), только косвенно связанные с консортом-детерминантом, и расчленяя консорцию на три круга - концентра (I, II, III). При этом консорты II и III концентров непосредственно связаны только с консортами предыдущего концентра, но не с консортом-детерминантом.
По-видимому, лучше сохранить то понятие консорции, которое предложили В.Н. Беклемишев и Л.Г. Раменский, как особой группы организмов в биоценозе (и соответственно в фитоценозе), связанных прямыми, непосредственными отношениями ее компонентов с детерминантным консортом. А.А. Корчагин (1976) справедливо замечает, что "... если встать на путь 'выделения в консорции трех концентров, то с тем же правом можно и дальше увеличивать число концентров (IV, V, VI и т.д.), в которых компоненты каждого последующего концентра так или иначе связаны с компонентами каждого предыдущего…; в результате представление о консорции станет безграничным".
В заключение следует отметить, что консорция представляет собой функциональную систему, компоненты которой не только связаны с центральным консортом-продуцентом, но и характеризуются сопряженной эволюцией и выработкой у них приспособлений друг к другу, что, между прочим, свидетельствует о большом значении коадаптации видов растений и животных в процессе исторического развития фитоценозов и биоценозов.
[1] В зарубежной литературе для обозначения данного показателя используется термин "видовое разнообразие", имея в виду один из его вариантов, а именно, альфа-разнообразие.
9Заметим, что здесь речь идет лишь о макрофитных синузиях. Если учитывать также микрофитные напочвенные и внутрипочвенные синузии, то окажется, что в природе нет односинузиальных фитоценозов.

