




Рассмотрев две самые важные сенсорные системы, опишем вкратце проприоцепцию, т.е. сенсорную систему, которая доставляет информацию о положении и движениях тела и его различных частей. Как всем известно, о положении и движениях головы, конечностей и всего тела мы обычно узнаём без помощи зрения. Даже при закрытых глазах мы знаем, вытянута или согнута у нас рука или нога, Повернута голова вперед или вбок, стоим ли мы прямо. Рецепторы этой сенсорной системы находятся в мышцах и суставных сумках, а также в вестибулярном аппарате внутреннего уха. Эти рецепторы получили название проприоцепторов; это значит, что они информируют нас о состоянии нашего собственного тела (от лат. proprius - собственный). Рецепторы в коже, о которых речь пойдет позднее, тоже участвуют в сигнализации об изменениях в положении тела; трудно отделить осязание и чувствительность к давлению от проприоцепции.
Не все элементы статической и кинетической информации (т.е. информации о положении и движениях тела) достигают большого мозга. Генерируемые в проприоцепторах импульсы могут активировать врожденные автоматизмы, приводящие к рефлекторной деятельности, которая регулируется не корой, а продолговатым или средним мозгом; иногда эти импульсы даже прямо проводятся к передним рогам спинного мозга. Некоторые движения, связанные с прямостоянием, совершаются автоматически. Реакции, связанные с тонким регулированием положения тела и головы, доходят до сознания с задержкой или же не доходят вовсе. Таким образом, некоторые процессы, происходящие в проприоцептивном анализаторе, остаются неосознанными, подобно переработке информации в висцеральном сенсорном аппарате, о котором речь пойдет позднее.
Строение и кодирующая активность проприоцепторов.
Проприоцеторы, как уже говорилось, расположены в скелетных мышцах, суставных сумках и внутреннем ухе.
Мышечные веретена и тельца Гольджи. Проприоцепторы, находящиеся в мышцах, заключены в веретенах - группах особых мышечных волокон длиной 3-4 мм, окруженных капсулой. Вокруг таких волокон обвиваются аннулоспиральные рецепторы. При растяжении мышцы импульсы от рецепторов направляются к нейронам в сенсорном ганглии, расположенном около спинного мозга. В мышечном веретене есть и другие, менее важные рецепторы В сухожилиях в месте их соединения с мышцами находятся рецепторы еще одного типа - тельца Гольджи. Импульсы в них возникают, когда мышца растянута или сокращена. Волокна от телец Гольджи тоже идут к сенсорному ганглию, лежащему около спинного мозга. Порог раздражения у мышечных веретен гораздо ниже, чем у сухожильных телец Гольджи. Для возбуждения этих последних требуется чрезвычайно сильное растяжение или сокращение. В чувствительных аннулоспиральных рецепторах импульсы возникают только в ответ на растяжение мышцы, в которой они заключены. Импульсы, как правило, направляются самым коротким сенсорным путем к спинному мозгу и возбуждают так называемый рефлекс на растяжение. Функция этой системы состоит в том, чтобы мышечные волокна принимали и сохраняли такую длину, какая требуется для данного положения тела. Стимулом для этого рефлекса служит земное притяжение, которое нужно преодолевать для поддержания нужной позы. Эти рефлексы называют также антигравитационными спинальными рефлексами. Импульсы не доходят до головного мозга, и рефлексы сохраняются у экспериментальных животных, лишенных всех нервных структур выше спинного мозга. Второй путь, по которому передаются проприоцептивные импульсы, несущие важную статическую и кинетическую информацию, идет к большому мозгу и мозжечку.
Лабиринт. Лабиринт, или вестибулярный аппарат внутреннего уха, является вторым важным проприоцептором. Он структурно и генетически не зависит от улитки, хотя лежит во внутреннем ухе непосредственно над слуховым аппаратом. Вестибулярный аппарат состоит из отолитовых органов (круглого и овального мешочков) и трех полукружных каналов, которые расположены в трех взаимно перпендикулярных плоскостях. Как и улитка, полукружные каналы заполнены эндолимфой. Рецепторными клетками, так же как и в кортиевом органе, являются волосковые клетки, погруженные в студенистую массу. Группы волосковых клеток найдены как в ампулах полукружных каналов, так и в отолитовых органах. Таким образом, каждый вестибулярный аппарат включает пять рецепторных систем. Волосковые клетки круглого и овального мешочков служат для получения информации о статическом положении головы. В студенистую массу погружены известковые частицы (отолиты), на которые воздействует сила тяжести. Порог раздражения этих рецепторов очень низок, поэтому импульсы от них постоянно идут в продолговатый мозг. Нервные окончания у основания волосковых клеток реагируют, когда известковые частички тянут волоски. Рецепторы полукружных каналов (волосковые клетки на кристах) воспринимают ускорение- положительное или отрицательное - движения головы. Таким образом, пока голова неподвижна, о ее положении сигнализируют в центр отолитовые органы, а о движениях головы сообщают рецепторы полукружных каналов. Порог раздражения полукружных каналов, реагирующих на изменения тока жидкости в их полости, выше, чем у отолитовых органов. Если голова неподвижна или движется с равномерной скоростью, эти рецепторы не посылают сигналов.
Не вся информация, передаваемая вестибулярным аппаратом, достигает коры. Часть ее доходит только до продолговатого или среднего мозга, приводя в действие сложный, но бессознательный автоматизм, регулирующий положение тела.
Корковое представительство. Проекционные зоны проприоцепторов в коре изучались методом вызванных потенциалов. Как выяснилось, проприоцепторы имеют свое представительство главным образом в мозжечке, который служит важным центром координации движений, но группы клеток, отвечающих на стимуляцию различных проприоцепторов (в мышцах или в вестибулярном аппарате), найдены также в соматосенсорной области коры (поля 1, 2 и 3 по Бродману). Поскольку нейроны в соматосенсорной коре расположены в соответствии с топографией тела (представительство головы и верхней части туловища находится в нижнем конце, близ сильвиевой борозды, а представительство ног - наверху), различные группы мышц тоже представлены в соответствующих частях соматосенсорной коры. Вестибулярный аппарат проецируется на представительство головы. Подобно зрительным и слуховым импульсам, сигналы, посылаемые проприоцепторами, тоже имеют станцию переключения в таламусе. Таламическое ядро n. ventralis posterior служит промежуточной станцией для всех кожных, мышечных и висцеральных сенсорных путей.
Таким образом, проприоцептивные импульсы идут по сложному, самовозбуждающемуся кольцевому пути, который включает сенсорные и моторные зоны коры, определенные группы нейронов в стволе мозга (анатомы назвали их базальными ганглиями), упомянутые выше нейроны таламуса и мозжечковые структуры. Эту реверберационную систему, которая регулирует позу и координирует движения, неврологи называют экстрапирамидной системой. В то время как пирамидный тракт, идущий от моторной коры к мотонейронам спинного мозга, состоит всего из двух или трех нейронов и осуществляет точные, запланированные произвольные движения, экстрапирамидный тракт управляет бессознательными, грубыми движениями. Рецепторами, снабжающими эту систему информацией, служат проприоцепторы, о которых шла речь выше.
Декодирование в коре информации о положении тела
Маунткасл и его сотрудники показали, что о положении и движениях конечностей мозгу сообщают проприоцепторы мышц, сухожилий и суставных сумок. В экспериментах прослеживалось движение периферических импульсов. Было установлено, что все сенсорные волокна, участвующие в регуляции движений конечностей, обладают угловым порогом возбуждения. Когда сгибание или выпрямление сустава достигает порогового угла, возникают импульсы, идущие к центру.
"Односторонняя" и "двусторонняя" реакция. Одни волокна обнаруживают "одностороннюю" реакцию: импульсация в них достигает наибольшей частоты при максимальном разгибании или сгибании конечности. Другим волокнам свойственна "двусторонняя" реакция: в этом случае частота импульсов достигает максимума при каком-то промежуточном положении конечности и снижается как при разгибании, так и при сгибании. Таким образом, мозг получает информацию и об угловом положении сустава, и о направлении движения. Об импульсации, поступающей ко второму нейрону ствола мозга, мало что известно.
Маунткасл получил очень интересные данные о декодировании импульсации в таламусе обезьяны. Здесь нейроны, как правило, дают "одностороннюю" реакцию, и их угловые пороги в 4 раза выше, чем у рецепторных волокон. Например, если максимальная частота импульсов в рецепторе возникает при разгибании сустава на 10° и на один таламическнй нейрон конвергируют четыре таких волокна, то он выдает максимальную импульсацию при 40°. В одних группах клеток при сгибании частота растет, в других она падает; в-третьих такого же рода реакцию может вызывать разгибание.
Отношение между углом сгибания сустава и частотой импульсации выражалась уравнением
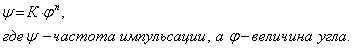
Психофизика и электрофизиология. Приведенное выше экспоненциальное уравнение идентично уравнению для психофизического закона Фехнера, видоизмененного Стивенсом. Это хороший пример совпадения психофизических и электрофизиологических данных, т.е. субъективного ощущения и результатов объективной регистрации импульсов, которые свидетельствуют о линейном преобразовании протекающих в мозгу процессов в информационной системе стимул-ощущение от рецепции стимула до словесного отчета.
Гамма-эфферентный механизм: центральный контроль мышечных рецепторов
Гамма-эфферентный механизм - это, пожалуй, самый известный и наиболее изученный пример центрального контроля рецепторной функции. Волокна этого эфферентного пути идут к мышечным веретенам прямо от спинного мозга и проводят импульсы от продолговатого мозга и даже от центров еще более высокого уровня. Эти эфферентные волокна названы гамма-волокнами из-за их размеров (класс волокон AY). Тела этих нейронов, так же как альфа-нейроны эффекторных мышечных нервов, находятся в спинном мозгу. Функция гамма-эфферентного механизма состоит в приспособлении сокращения мышечных волокон в веретене к данному состоянию скелетной мышцы. Импульсация гамма-эфферентных волокон, как правило, снижает порог возбуждения аннулоспиральных рецепторов. Этот механизм играет важную роль в поддержании определенного положения тела.
Гамма-эфферентный механизм составляет часть саморегулирующейся системы. Когда тело или какая-либо часть его принимает определенное положение, растяжению соответствующих мышц предшествует приход гамма-эфферентной импульсации, регулирующей частоту импульсов, порождаемых аннуло-спиральными рецепторами. Затем центрально управляемые моторные нервы вызывают сокращение соответствующих мышц, причем активность этих нервов контролируется рефлек-торно гамма-эфферентами. Таким образом, эта система включает один афферентный и два эфферентных пути (ее называют также гамма-петлей).

