




Особенности строения белков-рецепторов для стероидов.
Наиболее изученным является рецептор для гормонов коры надпочечников - глюкокортикостероидов(ГКС). В этом белке имеется три функциональных участка:
1 - для связывания с гормоном (С-концевой)
2 - для связывания с ДНК (центральный)
3 - антигенный участок, одновременно способный модулировать функцию промотора в процессе транскрипции (N-концевой).
Функции каждого участка такого рецептора ясны из их названий. Очевидно, что такое строение рецептора для стероидов позволяет им влиять на скорость транскрипции в клетке. Это подтверждается тем, что под действием стероидных гормонов избирательно стимулируется (или тормозится) биосинтез некоторых белков в клетке. В этом случае наблюдается ускорение (или замедление) образования мРНК. В результате изменяется количество синтезируемых молекул определенных белков (часто - ферментов) и меняется скорость метаболических процессов.
БИОСИНТЕЗ и СЕКРЕЦИЯ ГОРМОНОВ РАЗЛИЧНОГО СТРОЕНИЯ
Белково-пептидные гормоны.
В процессе образования белковых и пептидных гормонов в клетках эндокринных желез происходит образование полипептида, не обладающего гормональной активностью. Но такая молекула в своем составе имеет фрагмент(ы), содержащий(е) аминокислотную последовательность данного гормона. Такая белковая молекула называется пре-про-гормоном и имеет в своем составе (обычно на N-конце) структуру, которая называется лидерной или сигнальной последовательностью (пре-). Эта структура представлена гидрофобными радикалами и нужна для прохождения этой молекулы от рибосом через липидные слои мембран внутрь цистерн эндоплазматического ретикулума (ЭПР). При этом, во время перехода молекулы через мембрану в результате ограниченного протеолиза лидерная (пре-) последовательность отщепляется и внутри ЭПР оказывается прогормон. Затем через систему ЭПР прогормон транспортируется в комплекс Гольджи и здесь заканчивается созревание гормона. Вновь в результате гидролиза под действием специфических протеиназ отщепляется оставшийся (N-концевой) фрагмент (про-участок). Образованная молекула гормона, обладающая специфической биологической активностью поступает в секреторные пузырьки и накапливается до момента секреции.
При синтезе гормонов из числа сложных белков гликопротеинов (например, фолликулостимулирующего (ФСГ) или тиреотропного (ТТГ) гормонов гипофиза) в процессе созревания происходит включение углеводного компонента в структуру гормона.
Может происходить и внерибосомальный синтез. Так синтезируется трипептид тиролиберин (гормон гипоталамуса).
Гормоны - производные аминокислот
Из тирозина синтезируются гормоны мозгового слоя надпочечников АДРЕНАЛИН и НОРАДРЕНАЛИН, а также ЙОДСОДЕРЖАЩИЕ ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ. В ходе синтеза адреналина и норадреналина тирозин подвергается гидроксилированию, декарбоксилированию и метилированию с участием активной формы аминокислоты метионина.
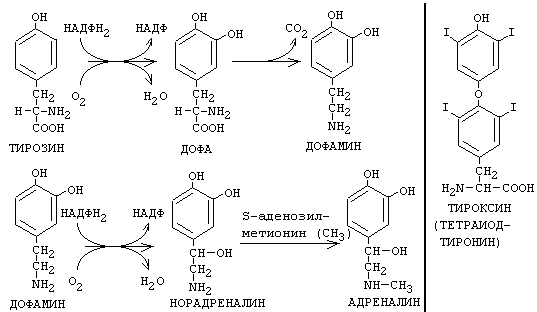
В щитовидной железе происходит синтез йодсодержащих гормонов трийодтиронина и тироксина (тетрайодтиронина). В ходе синтеза происходит йодирование фенольной группы тирозина. Особый интерес представляет метаболизм иода в щитовидной железе. Молекула гликопротеина тиреоглобулина (ТГ) имеет молекулярную массу более 650 кДа. При этом в составе молекулы ТГ около 10% массы - углеводы и до 1% - йод. Это зависит от количества иода в пище. В полипептиде ТГ - 115 остатков тирозина, которые иодируются окисленным с помощью специального фермента - тиреопероксидазы - йодом. Эта реакция называется органификацией йода и происходит в фолликулах щитовидной железы. В результате из остатков тирозина образуются моно- и ди-иодтирозин. Из них примерно 30% остатков в результате конденсации могутпревратитьться в три- и тетра- иодтиронины. Конденсация и иодирование идут с участием одного и того же фермента - тиреопероксидазы. Дальнейшее созревание гормонов щитовидной железы происходит в железистых клетках - ТГ поглощается клетками путем эндоцитоза и образуется вторичная лизосома в результате слияния лизосомы с поглощенным белком ТГ.
Протеолитические ферменты лизосом обеспечивают гидролиз ТГ и образование Т3 и Т4, которые выделяются во внеклеточное пространство. А моно- и дииодтирозин деиодируются с помощью специального фермента деиодиназы и иод повторно может подвергаться органификации. Для синтеза тиреоидных гормонов характерным является механизм торможения секреции по типу отрицательной обратной связи (Т3 и Т4 угнетают выделение ТТГ).
Стероидные гормоны.
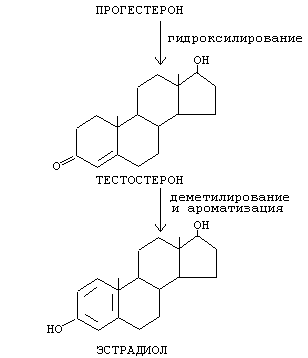
Стероидные гормоны синтезируются из холестерина (27 углеродных атомов), а холестерин синтезируется из ацетил-КоА.

|
Холестерин превращается в стероидные гормоны в результате следующих реакций:
- отщепление бокового радикала
- образование дополнительных боковых радикалов в результате реакции гидроксилирования с помощью специальных ферментов монооксигеназ (гидроксилаз) - чаще всего в 11-м, 17-м, и 21-м положениях (иногда в 18-м). На первом этапе синтеза стероидных гормонов сначала образуются предшественники (прегненолон и прогестерон), а затем другие гормоны (кортизол, альдостерон, половые гормоны). Из кортикостероидов могут образоваться альдостерон, минералокортикоиды.
СЕКРЕЦИЯ ГОРМОНОВ.
Регулируется со стороны ЦНС. Синтезированные гормоны накапливаются в секреторных гранулах. Под действием нервных импульсов или под влиянием сигналов из других эндокринных желез (тропные гормоны) в результате экзоцитоза происходит дегрануляция и выход гормона в кровь.
Механизмы регуляции в целом были представлены в схеме механизма реализации эндокринной функции.
ТРАНСПОРТ ГОРМОНОВ.
Транспорт гормонов определяется их растворимостью. Гормоны, имеющие гидрофильную природу (например, белково-пептидные гормоны) обычно транспортируются кровью в свободном виде. Стероидные гормоны, йодсодержащие гормоны щитовидной железы транспортируются в виде комплексов с белками плазмы крови. Это могут быть специфические транспортные белки (транспортные низкомолекулярные глобулины, тироксинсвязывающий белок; транспортирующий кортикостероиды белок транскортин) и неспецифический транспорт (альбумины).
Уже говорилось о том, что концентрация гормонов в кровяном русле очень низка. И может меняться в соответствии с физиологическим состоянием организма. При снижении содержания отдельных гормонов развивается состояние, характеризуемое как гипофункция соответствующей железы. И, наоборот, повышение содержания гормона - это гиперфункция.
Постоянство концентрации гормонов в крови обеспечивается также процессами катаболизма гормонов.
КАТАБОЛИЗМ ГОРМОНОВ.
Белково-пептидные гормоны подвергаются протеолизу, распадаются до отдельных аминокислот. Эти аминокислоты вступают дальше в реакции дезаминирования, декарбоксилирования, трансаминирования и распадаются до до конечных продуктов: NH3, CO2 и Н2О.
Гормоны - производные аминокислот подвергаются окислительному дезаминированию и дальнейшему окислению до СО2 и Н2О. Стероидные гормоны распадаются иначе. В организме нет ферментных систем, которые обеспечивали бы их распад. Что же происходит при их катаболизме?
В основном происходит модификация боковых радикалов. Вводятся дополнительные гидроксильные группы. Гормоны становятся более гидрофильными. Образуются молекулы, представляющие собой структуру стерана, у которого в 17-м положении находится кетогруппа. В таком виде продукты катаболизма стероидных половых гормонов выводятся с мочой и называются 17-КЕТОСТЕРОИДЫ. Определение их количества в моче и крови показывает содержание в организме половых гормонов.

