




Гидатофиты – это водные растения, целиком или почти целиком погруженные в воду. Среди них – цветковые, которые вторично перешли к водному образу жизни (элодея, рдесты, водяные лютики, валлиснерия, уруть и др.). Вынутые из воды, эти растения быстро высыхают и погибают. У них редуцированы устьица и нет кутикулы. Транспирация у таких растений отсутствует, а вода выделяется через особые клетки – гидатоды.
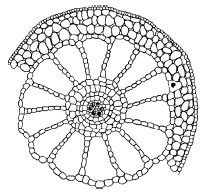
Рис. 32. Поперечный срез стебля урути Myriophyllum verticillatum (по Т. К. Горышиной, 1979)
Листовые пластинки у гидатофитов, как правило, тонкие, без дифференцировки мезофилла, часто рассеченные, что способствует более полному использованию ослабленного в воде солнечного света и усвоению СО2. Нередко выражена разнолистность – гетерофиллия; у многих видов есть плавающие листья, имеющие световую структуру. Поддерживаемые водой побеги часто не имеют механических тканей, в них хорошо развита аэренхима (рис. 32).
Корневая система цветковых гидатофитов сильно редуцирована, иногда отсутствует совсем или утратила свои основные функции (у рясок). Поглощение воды и минеральных солей происходит всей поверхностью тела. Цветоносные побеги, как правило, выносят цветки над водой (реже опыление совершается в воде), а после опыления побеги снова могут погружаться, и созревание плодов происходит под водой (валлиснерия, элодея, рдесты и др.).
Гидрофиты – это растения наземно-водные, частично погруженные в воду, растущие по берегам водоемов, на мелководьях, на болотах. Встречаются в районах с самыми разными климатическими условиями. К ним можно отнести тростник обыкновенный, частуху подорожниковую, вахту трехлистную, калужницу болотную и другие виды. У них лучше, чем у гидатофитов, развиты проводящие и механические ткани. Хорошо выражена аэренхима. В аридных районах при сильной инсоляции их листья имеют световую структуру. У гидрофитов есть эпидерма с устьицами, интенсивность транспирации очень высока, и они могут расти только при постоянном интенсивном поглощении воды.
Гигрофиты – наземные растения, живущие в условиях повышенной влажности воздуха и часто на влажных почвах. Среди них различают теневые и световые. Теневые гигрофиты – это растения нижних ярусов сырых лесов в разных климатических зонах (недотрога, цирцея альпийская, бодяк огородный, многие тропические травы и т. п.). Из-за высокой влажности воздуха у них может быть затруднена транспирация, поэтому для улучшения водного обмена на листьях развиваются гидатоды, или водяные устьица, выделяющие капельно-жидкую воду. Листья часто тонкие, с теневой структурой, со слабо развитой кутикулой, содержат много свободной и малосвязанной воды. Обводненность тканей достигает 80 % и более. При наступлении даже непродолжительной и несильной засухи в тканях создается отрицательный водный баланс, растения завядают и могут погибнуть.
К световым гигрофитам относятся виды открытых местообитаний, растущие на постоянно влажных почвах и во влажном воздухе (папирус, рис, сердечники, подмаренник болотный, росянка и др.). Переходные группы – мезогигрофиты и гигромезофиты.
Мезофиты могут переносить непродолжительную и не очень сильную засуху. Это растения, произрастающие при среднем увлажнении, умеренно теплом режиме и достаточно хорошей обеспеченности минеральным питанием. К мезофитам можно отнести вечнозеленые деревья верхних ярусов тропических лесов, листопадные деревья саванн, древесные породы влажных вечнозеленых субтропических лесов, летнезеленые лиственные породы лесов умеренного пояса, кустарники подлеска, травянистые растения дубравного широкотравья, растения заливных и не слишком сухих суходольных лугов, пустынные эфемеры и эфемероиды, многие сорные и большинство культурных растений. Из приведенного перечня видно, что группа мезофитов очень обширна и неоднородна. По способности регулировать свой водный обмен одни приближаются к гигрофитам (мезогигрофиты), другие – к засухоустойчивым формам (мезоксерофиты).
Ксерофиты растут в местах с недостаточным увлажнением и имеют приспособления, позволяющие добывать воду при ее недостатке, ограничивать испарение воды или запасать ее на время засухи. Ксерофиты лучше, чем все другие растения, способны регулировать водный обмен, поэтому и во время продолжительной засухи остаются в активном состоянии. Это растения пустынь, степей, жестколистных вечнозеленых лесов и кустарниковых зарослей, песчаных дюн.
Ксерофиты подразделяются на два основных типа: суккуленты и склерофиты.
Суккуленты – сочные растения с сильно развитой водозапасающей паренхимой в разных органах. Стеблевые суккуленты – кактусы, стапелии, кактусовидные молочаи; листовые суккуленты – алоэ, агавы, мезембриантемумы, молодило, очитки; корневые суккуленты – аспарагус. В пустынях Центральной Америки и Южной Африки суккуленты могут определять облик ландшафта (рис. 33).

Рис. 33. Суккулентное растение – древовидный кактус из пустыни Аризона
Листья, а в случае их редукции стебли суккулентов имеют толстую кутикулу, часто мощный восковой налет или густое опушение. Устьица погруженные, открываются в щель, где задерживаются водяные пары.
Днем они закрыты. Это помогает суккулентам сберегать накопленную влагу, но зато ухудшает газообмен, затрудняет поступление СО2 внутрь растения. Поэтому многие суккуленты из семейств лилейных, бромелиевых, кактусовых, толстянковых ночью при открытых устьицах поглощают СО2, который только на следующий день перерабатывают в процессе фотосинтеза. Поглощенный СО2 переводится в малат. Кроме того, при дыхании ночью углеводы разлагаются не до углекислого газа, а до органических кислот, которые отводятся в клеточный сок. Днем на свету малат и другие органические кислоты расщепляются с выделением СО2, который используется в процессе фотосинтеза. Таким образом, крупные вакуоли с клеточным соком запасают не только воду, но и СО2. Так как у суккулентов ночная фиксация углекислоты и переработка ее днем в ходе фотосинтеза разделены во времени, они обеспечивают себя углеродом, не подвергаясь риску чрезмерной потери воды, но масштабы поступления углекислого газа при таком способе невелики, и растут суккуленты медленно.
Осмотическое давление клеточного сока суккулентов мало – всего 3 · 105 – 8 · 105 Па (3–8 атм), они развивают небольшую сосущую силу и способны всасывать воду лишь атмосферных осадков, просочившихся в верхний слой почвы. Корневая система их неглубокая, но сильно распростертая, что особенно характерно для кактусов.
Склерофиты – это растения, наоборот, сухие на вид, часто с узкими и мелкими листьями, иногда свернутыми в трубочку. Листья могут быть также рассеченными, покрытыми волосками или восковым налетом. Хорошо развита склеренхима, поэтому растения без вредных последствий могут терять до 25 % влаги не завядая. В клетках преобладает связанная вода. Сосущая сила корней до нескольких десятков атмосфер, что позволяет успешно добывать воду из почвы. При недостатке воды резко снижают транспирацию. Склерофиты можно подразделить на две группы: эуксерофитов и стипаксерофитов.
К эуксерофитам относятся многие степные растения с розеточными и полурозеточными, сильно опушенными побегами, полукустарнички, некоторые злаки, полынь холодная, эдельвейс эдельвейсовидный и др. Наибольшую биомассу эти растения создают в период, благоприятный для вегетации, а в жару уровень обменных процессов у них очень низок.
Стипаксерофиты – это группа узколистных дерновинных злаков (ковыли, тонконоги, типчак и др.). Характеризуются низкой транспирацией в засушливый период и могут переносить особенно сильное обезвоживание тканей. Свернутые в трубочку листья имеют внутри влажную камеру. Транспирация идет через погруженные в бороздки устьица внутрь этой камеры, что снижает потери влаги (рис. 34).
А, Б
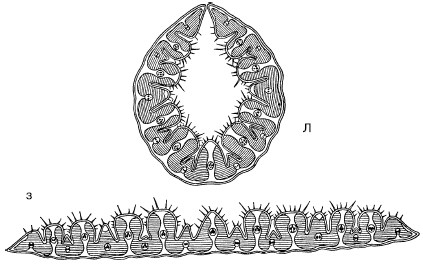
Рис. 34. Поперечный срез листа ковыля Stipa capillata (по A. Кернеру, 1896): A – при засухе (лист свернут); Б – во влажную погоду (пластинка листа развернута)
Кроме названных экологических групп растений, выделяют еще целый ряд смешанных или промежуточных типов.
Различные пути регуляции водообмена позволили растениям заселить самые различные по экологическим условиям участки суши. Многообразие приспособлений лежит, таким образом, в основе распространения растений по поверхности земли, где дефицит влаги является одной из главных проблем экологических адаптаций.

