









 Z-стратегия
Z-стратегия
г-стратегия

 Конкурентоспособность за Конкурентоспособность за счет быстрой реакции на уве-
Конкурентоспособность за Конкурентоспособность за счет быстрой реакции на уве-
счет низкой пороговой кон- личение доступности ресурсов в благоприятные периоды,
центрации ресурсов Периоды с низкой доступностью ресурсов переживают в:

 толерантной стадии
толерантной стадии
латентной стадии



 ПАТИЕНТЫ
ПАТИЕНТЫ
1.  Произрастание в небла
Произрастание в небла
гоприятных условиях
(абиотический и биоти
ческий стресс)
2. Отрицательная реакция
на нарушения
3. Обычно невысокое ре
продуктивное усилие
4. Невысокий запас жизне
способных семян в почве
[ ВИОЛЕНТЫ)
1. Большая биомасса в благоприятных условиях
2. Отрицательная реакция
на нарушения
3. Высокое репродуктивное
усилие при крупных раз
мерах семян
4. Практическое отсутствие
семенного запаса в почве
| ЭКСПЛЕРЕНТЫ |
1. Произрастание в благоприятных условиях
2. Положительная реакция
на нарушения
3. Высокое репродуктивное
усилие при мелких раз
мерах семян
4. Высокий запас жизне
способных семян в почве
Факторы, не зависящие от плотности населения. К этой группе2 относится комплекс абиотических факторов, которые в своем воздействии на животных реализуются через составляющие климата и погоды. Биологическое влияние этих факторов характеризуется тем, что они действуют на уровне организма и именно поэтому эффект их воздействия не связан с такими специфическими популяционными параметрами, как численность и плотность
 Это не единственная попытка классификации экологических факторов в связи с их ролью в динамике численности. Так, А.С. Мончадский (1962) делил факторы на первично-периодические (солнечная радиация, температура), вторично-периодические (атмосферная влажность, урожай кормовых растений, внутривидовые отношения) и изменяющиеся незакономерно (осадки, ветер, паразиты, хищники, болезни). Из них ведущую роль в формировании циклов численности он отводил вторично-периодическим факторам, поскольку к первично-периодическим эволюционно вырабатываются эффективные адаптации, а непериодические факторы влияют на численность случайным образом.
Это не единственная попытка классификации экологических факторов в связи с их ролью в динамике численности. Так, А.С. Мончадский (1962) делил факторы на первично-периодические (солнечная радиация, температура), вторично-периодические (атмосферная влажность, урожай кормовых растений, внутривидовые отношения) и изменяющиеся незакономерно (осадки, ветер, паразиты, хищники, болезни). Из них ведущую роль в формировании циклов численности он отводил вторично-периодическим факторам, поскольку к первично-периодическим эволюционно вырабатываются эффективные адаптации, а непериодические факторы влияют на численность случайным образом.
Ч. Элтон называл их «факторами среды» (Ch. Elton, 1927), а Н.П. Наумов (1953) — «экзогенными факторами».
населения. Действие этих факторов односторонне: организмы могут к ним приспосабливаться, но не в состоянии оказать на них обратное влияние.
Эффект воздействия климатических факторов на уровень численности и направленность ее изменения реализуется в первую очередь через изменения смертности, возрастающей по мере отклонения силы воздействующего фактора от оптимальной величины. При этом уровень смертности и выживания определяется только силой воздействующего фактора с учетом адаптивных возможностей организма и некоторых характеристик среды: наличие убежищ с более благоприятными условиями, смягчающее действие попутных факторов и т. п. Уровень численности (плотности населения) популяции не определяет общей направленности ее изменений, а в ряде случаев и конечного результата. Так, если зимние условия отличаются необычно низкими температурами при небольшой толщине снежного покрова, уровень весенней численности мелких грызунов окажется крайне низким даже в том случае, если к началу зимы популяция достигла высокой численности и плотности населения. Напротив, если зимние условия благоприятны, относительная величина сезонного падения численности будет невелика независимо от плотности населения осенью. Аналогичная закономерность характерна для лесных куриных птиц, спасающихся от жестоких морозов в снежных лунках.
При некоторых условиях факторы этой группы могут действовать и путем изменения плодовитости, главным образом косвенно, через изменения кормовых условий. Например, комплекс абиотических факторов, благоприятный для вегетации растений, способствует лучшему размножению животных-фитофагов. Напротив, полный неурожай или недоступность корма увеличивают независимую от плотности гибель. Известно, в частности, явление массового падежа копытных при длительной гололедице.
Наиболее полно и логично эта система связей сформулирована в трофоклиматической теории И.Я. Полякова (1949, 1950, 1976). Однако в полной мере эта концепция применима лишь при условии, что динамика пищевых ресурсов в слабой степени зависит от пресса потребителей. В более общей форме реализация взаимоотношений потребителя и пищи относится к сфере биоценотических процессвв и зависит от плотности населения взаимодействующих видов (см. ниже).
Связь влияния абиотических факторов со структурой популяции может выражаться в избирательно повышенной смертности определенных демографических групп животных в популяциях (например, молодняка, мигрантов и др.); на основе изменения демографической структуры популяции может, как вторичный эффект, меняться и уровень воспроизведения. Показано, например, что в популяциях белки Sciurus vulgaris преобладание молодых животньк, отличающихся меньшей плодовитостью, снижает общую интенсивность размноже-

|
факторов в формировании характера динамики численности, метеорологические условия относятся к категории модифицирующих факторов.
Факторы, зависящие от плотности населения. Эта группа факторов («факторы авторегуляции» или эндогенные факторы) включает влияние на уровень и динамику численности данного вида его пищи, хищников, возбудителей болезней и др. Характер влияния зависящих от плотности факторов принципиально отличается от факторов, рассмотренных выше: действуя на численность популяций других видов, они сами испытывают влияние с их стороны. Таким образом, в этом случае правильнее говорить о взаимодействиях популяций разных видов в составе биоценоза, выступающих в роли биоценотического регулятора численности обоих видов. Регуляторный эффект такого рода отношений зависит от плотности населения взаимодействующих популяций.
Г.А Викторов (1965, 1967), создавший объективно обоснованную концепцию факторов динамики численности, относит биотические взаимодействия к категории регулирующих факторов (механизмов) именно на основе способности популяций реагировать как на изменения собственной плотности, так и плотности популяций других видов, с которыми они связаны трофическими или иными взаимоотношениями. Регуляция в этом случае осуществляется по кибернетическому принципу обратных связей. Этот принцип исходно содержит в себе предпосылки колебательного процесса, так как регулирующий механизм управляется отклонениями самой регулируемой величины (в данном случае плотности населения). Эффект действия проявляется с некоторым замедлением; в результате плотность популяции проявляет закономерные колебания вокруг оптимального в данных условиях уровня. Значение биоценотических взаимоотношений как фактора, формирующего циклы численности, подчеркивалось уже давно (Ch. Elton, 1927; A. Nicholson, 1933; A Nicholson, V. Bailey, 1935; С.А. Северцов, 1936,1941, 1942) и рассматривается как один из ведущих механизмов циклической динамики численности в современной экологии.
Одна из важных для формирования циклов численности форм биотических взаимоотношений — отношения потребителя и его пищи. В наиболее простом варианте роль пищи как фактора индукции циклов сводится к тому, что высокая обеспеченность пищей вызывает рост рождаемости и уменьшение смертности в популяции потребителей. В результате их численность нарастает, что ведет к усиленному выеданию пищи и соответственно к снижению ее численности (биомассы). Последнее же означает ухудшение условий жизни потребителя, падение рождаемости, увеличение смертности и снижение численности. В результате снижается пресс на популяцию кормовых организмов, повышается ее численность (биомасса), что способствует росту численности потребителя, и цикл начинается снова. Классический пример такого рода взаимоотношений — так называемые «лемминговые пик-
лы», сопровождающиеся катастрофическими нарушениями тундровой растительности, массовыми ненаправленными миграциями и гибелью животных. Особенно отчетливо такие циклы выражены у норвежского Lemmus lemmus и бурого L. trimucronatis леммингов.
Но даже в этом, казалось бы, простом случае механизм формирования циклов численности оказывается более сложным, чем просто нехватка пищи и ее последующее восстановление. Период пика численности связан с увеличением агрессии, возникновением стресса в популяции и т. п. Соответственно снижение уровня репродукции идет через механизмы популяционной регуляции (Т. В. Кошкина, 1962; L. Коек etal., 1969, Ф.Б. Чернявский, 1975; АВ. Ткачев, 1980; Ф.Б. Чернявский, А. В. Ткачев, 1982). Выедание растительности ведет не только к недостатку пищи, но и к изменению ее качества (недостаток фосфора, кальция, белка и пр.), снижению защитных функций растительного покрова и соответствующему увеличению пресса хищников и т. п. Таким образом, и увеличение смертности имеет более сложный характер, чем простое голодание (F. Pitelka, 1958; A. Schultz, 1962).
Сложный характер влияния пищи на изменения численности показан и в других случаях. Так, у грызунов ухудшение питания вызывает задержку полового созревания, изменения пространственной структуры популяций и поведения животных (Н. Andreassen, R. Ims, 1990). У клариевого сома Clarias gariepinus нехватка пищи сказывается на замедлении роста молоди и индуцирует родственный каннибализм (Т. Hecht, S. Appelbaum, 1988). Наблюдения за популяциями моллюска блюдечко Patella granularis обнаружили связь уровня первичной продукции литоральных водорослей и ряда демографических параметров моллюсков (A. Bosnian, P. Hockey, 1988).
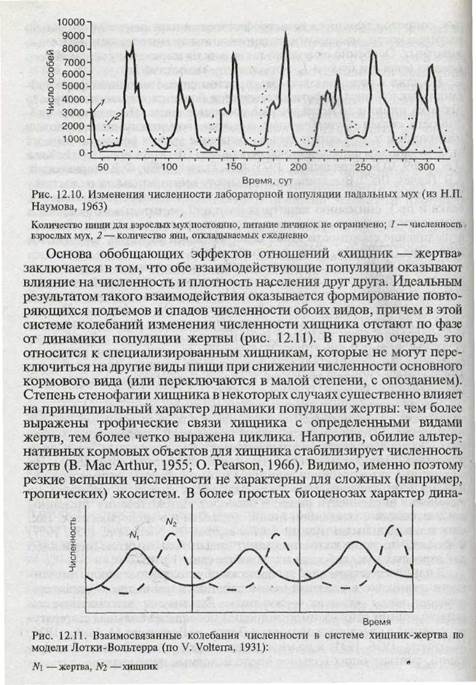
Тем не менее общее влияние обеспеченности пищей на изменения численности потребителей несомненно и показано данными многих наблюдений. Периодичность таких изменений численности может быть обусловлена цикличностью динамики кормового вида, зависящей от третьих факторов (например, по модели климатического цикла). Но в принципе автоколебания заложены в самой системе взаимоотношений на базе «лаг-эффекта» — запаздывания ответа потребителя на изменения численности пищи, и наоборот. Автоколебания численности, связанные с выеданием пищи, показаны и в экспериментах. Так, опыты с падальными мухами Lucilla cuprina (A. Nicholson, 1954, 1957) показали, что такие колебания могут вызываться лимитом пищи либо для взрослых мух, либо для их личинок (рис. 12.10).
В наиболее четком виде трофически обусловленные циклы численности возникают в условиях взаимодействия пары видов — взаимоотношения типа «хищник — жертва». Анализу и математической интерпретации этих взаимоотношений посвящена большая литература, начиная с классической модели Лотки — Вольтерра (A. Lotka, 1925; V. Volterra, 1926, 1931) и до многочисленных современных ее модификаций, учитывающих большое число исходных параметров.
|
мики и численности даже одного и того же вида может меняться, причем в относительно небольших географических масштабах. Установлено, например, что на севере Фенноскандии для полевок характерны циклические изменения численности, тогда как на юге региона численность относительно устойчива и циклы не формируются. Предполагается, что это связано с тем, что на юге хищники менее специализированы, поскольку более разнообразен видовой состав грызунов (L. Hansson, 1987). Анализ динамики популяций мелких грызунов в Норвегии за длительный (1871—1949) промежуток времени показал, что свойственная им цикличность не проявлялась в начале 1900-х годов; авторы связывают это с резким снижением хищничества — именно в эти годы шла программа по борьбе с хищниками (Н. Steen etal., 1990).
Однако примеров, точно соответствующих теоретическим расчетам динамики взаимодействующих видов в системе «хищник — жертва», при изучении естественных популяций практически не найдено. Даже прошедший через все учебники пример сопряженных колебаний численности американского беляка Lepus americanus и рыси Lynx lynx (no данным многолетних заготовок пушнины в Канаде) при корректном анализе и математической обработке оказался не соответствующим этой модели (М. Gilpin, 1973; М. Weinstein, 1977). Причина заключается не в ошибочности модели, а в том, что реальная динамика численности идет под влиянием многих факторов; взаимодействия хищников и жертв — лишь один из них. В целом многие авторы склонны считать хищничество важным фактором в динамике численности жертв, однако его роль в основном сводится либо к купированию пика численности, либо к задержке фазы ее роста.
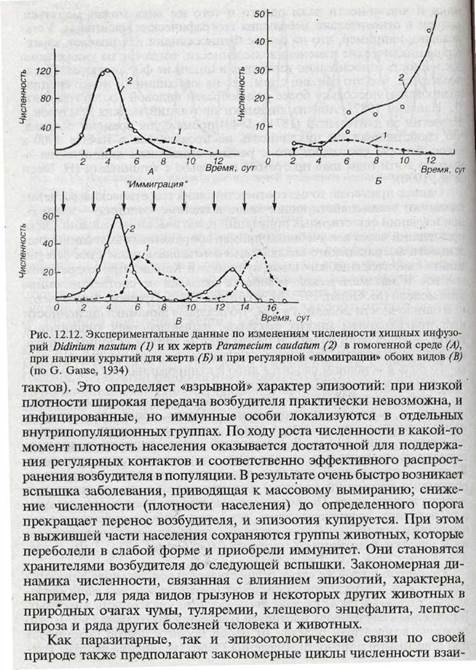
В классических экспериментах Г.Ф. Гаузе, поставленных с инфузориями в условиях, исключающих действие попутных факторов, воспроизвести повторяющиеся циклы в системе «хищник — жертва» не удалось (рис. 12.12). В однородной среде хищники полностью выедали своих жертв и вымирали сами; реализовался лишь один цикл. В среде, включающей укрытия для жертв, хищники вымирали, а популяция парамеций (жертвы) демонстрировала начало типичного логистического роста. Последовательные циклы обоих видов, сходные с расчетными, воспроизводились лишь при условии регулярного добавления в культуру особей обоих видов, что имитировало иммиграцию их в участки пониженной плотности (G. Gause, 1934; Г.Ф. Гаузе, А.А. Вйтт, 1934; Г.Ф. Гаузе, 1935).
Примерно по той же схеме осуществляется сопряженная динамика численности в паразитарных системах {взаимоотношения типа «паразит — хозяин»), а также влияние эпизоотопий на численность животных. В последнем случае роль плотности населения проявляется в том, что передача возбудителя болезни от одной особи к другой облегчается при повышении плотности населения (через увеличение частоты кон-
|
мосвязанных популяции, что подтверждается и математическим моделированием этих процессов. Имеются и наблюдения в природе, подтверждающие принципиальную возможность формирования закономерных циклов на базе рассмотренных типов биоценотических взаимоотношений. Однако примеров полной реализации таких циклов в природных условиях относительно немного; в большинстве случаев они осуществляются на протяжении сравнительного короткого промежутка времени, после чего следуют отклонения от классической схемы. Так, например, зимние вспышки туляремии обычно купируются весной, так как возбудитель не может долгое время существовать во внешней среде при высокой температуре. Кроме того, в биоценотических взаимодействиях природного очага болезни участвует комплекс видов и каждый из них подвержен действию многих факторов, влияющих на его численность и биологическую активность.
Все это, вместе взятое, привело к тому, что ряд исследователей скептически относятся к оценке регуляторнои роли биоценотических отношений, считая значение этих механизмов ненадежным и несовершенно зависящим от плотности. Полагают, что главная роль этой группы механизмов заключается лишь в «коррекции» отдельных фаз цикла изменений численности (V. Wynne-Edwards, 1962, 1966; Н.П. Наумов, 1963; А.А. Максимов, 1984). Вероятно, такая концепция исходит не из отрицания участия биоценотических связей в формировании циклов, а из сложного характера этого процесса, в котором невозможно выделить один ведущий фактор или даже небольшое их число.
Относительная роль отдельных факторов в комплексе, определяющем общий уровень численности и тенденции к ее изменениям, меняется в зависимости от условий местности и состояния популяции в разные сезоны и годы. Применение методов многофакторного анализа позволяет оценить эту роль в различных вариантах экосистем в разные годы. Многолетние данные по динамике популяций рыжей полевки в Карелии, например, показывают, что успешность перези-мовывания и интенсивность весеннего размножения в первую очередь связаны с демографической структурой популяции осенью; размножение и уровень численности в течение репродуктивного сезона определяются в основном внутрипопуляционными факторами, а в остальное время года —внешними воздействиями. При этом внешние факторы определяют верхний предел оптимальной плотности, а соответствие численности этим условиям регулируется внутрипопуляционными механизмами (Э.В. Ивантер и др., 1991).
Стационарные наблюдения за островной популяцией того же вида в Польше также показали, что размножение регулируется в первую очередь внутрипопуляционными механизмами, тогда как уровень выживания — главным образом внешними факторами, которые вызыва-

