




Ступінь покриття ґрунту окремими видами характеризує участь останніх у загальному синтезі органічної речовини з вуглекислого газу, води і зольних елементів у даному рослинному угрупованні. Тому види, які дають найбільше покриття, є домінуючими, а саме поняття про ступінь покриття замінюється поняттям про ступінь домінування.
Зімкнутість крон - це проективне покриття, створене проекціями без урахування просвітів крон дерев до поверхні ґрунту. Цей показник має велике значення при дослідженні лісових ценозів, оскільки від ступеня зімкнутості крон залежить світловий режим під пологом лісу.
Зімкнутість крон дає уявлення про густоту чи щільність деревостану. Чим вона більша, тим більші густота і щільність стояння рослин.
Проекція крони дерев не може бути ототожнена з площею світлокористування, оскільки площа її контуру не відповідає сумарній площі всіх листків крони, тобто площі світлокористування.
Бургер (1939), обстежуючи 35-річний ялиновий ліс, на площі 1 га росло 3148 дерев, сума проекцій крон яких не перевищувала 1 га, а сумарна поверхня всієї хвої становила понад 19 га. П.Б. Раскатов у дубовому 25-річному лісі визначив площу листків дуба, у якого в сумі вона становила понад 7 га. Площа листків конюшини на ділянці 1 га в 26 разів перевищує площу зайняту нею, а люцерни – в 85 разів.
Площа проективного покриття близька до площі світлокористування чи збігається з нею в трав'янистих розеткових та низькорослих рослин.
При вивченні всіх типів рослинності в геоботаніці користуються поняттям "проективна повнота, або загальне проективне покриття", і визначають цей показник у відсотках. Основними методами визначення проективного покриття є наступні.
Метод визначення проективного покриття
за допомогою сітки Л.Г. Раменського.
Сітка Л.Г. Раменського являє собою рамку з натягнутим на неї (вздовж і впоперек) дротом, з таким розрахунком, щоб площа однієї комірки становила 10 см2. Такі сітки бувають малі (розміром 2x5 см, великі 1x1 м з віконцем 10x10 см).
Проективну повноту покриву визначають, розглядаючи травостій крізь віконце сіточки. При цьому по можливості слід не звертати увагу на суцвіття. Малу сітку розмірами 2x5 см тримають приблизно на половині відстані міжоком і травостоєм, а велику сітку (вона значно зручніша) – на рівні травостою. Розглядаючи певне місце покриву, визначають, скільки комірок сітки (десятих часток віконця) припадає на плями (проекції рослин) і скільки – на вільні проміжки (крізь них видно ґрунт, воду мохи)
Для контролю користуються двома основними прийомами оцінки.

Сіточка для обліку повноти проекції (за Раменським)- А - віконце малої сітки натуральної величини; Б - зеркальна сіточка (за Понятовською)
Перший прийом полягає в тому, що уявно скупчують плямі (проекції) або вільні проміжки до одного кінця сіточки визначають, скільки комірок вони зайняли б при такому суцільному розміщенні. Якщо проекція рослин займає, наприклад, при цьому чотири комірки, то проективна повнота покриву становить 40%. Якщо вільні проміжки займаютьдві комірки, то проективна повнота покриву становить 80% і т.д.
Другий прийом полягає в тому, що подумки ділять сіточку впоперек на дві частини так, щоб плями (проекції) однієї частини заповнили вільні проміжки другої частини. Якщо, наприклад, достатньо виділити три комірки, щоб зробити суцільною ("залатати") проекцію в інших семи комірках, то повнота покриву дорівнює 70%.
Визначивши повноту покриву в кількох місцях обстежуваної Ділянки, враховують середню величину її. Якщо повнота покриву в окремих точках різко коливається, то площу обліку розбивають на частини за двома або трьома ступенями густоти і вираховують повноту покриву ділянки як середнє з цих двох або трьох величин.
Розглянемо такий приклад: третина облікової площі припадає на густі плями із середньою повнотою 80%; половина – на середньогустий травостій з повнотою 60% з розрахунку; а решта на – зріджений травостій, із середньою повнотою 30%. Тоді середня повнота всієї облікової площі становить 60%:
(0 3 х 80% + 0,5 х 60% + 0,2 х 30% =2 4 + 30 + 6 = 60%).
Такий прийом визначення повноти називають "двомірним".
Метод дзеркальної сіточки.
Дзеркальна сіточка являє собою картонну (або з іншого матеріалу) платівку, з віконцем, поділену дротиками на 10 рівних квадратів. Платівка рухливо скріплюється із дзеркальцем. Розглядаючи травостій крізь сіточку, розраховують повноту покриття так само, як за допомогою сіточки Л.Г. Раменського, а розглядаючи деревний ярус, відображений крізь віконце в дзеркальці, оцінюють його проективне покриття (зімкнутість крон).
Метод масштабної сіточки.
Для обрахунку відсотку покриття окремих видів у фітоценозі Л.Г. Раменський розробив метод масштабної проекційної сіточки, в якої розмір комірки становить 10%, 1% від 4 м2, тобто розмір найдрібніших комірок її 4 см2. Користуються масштабною сіточкою таким чином: над певною ділянкою травостою розтягують квадратну сітку площею 4 м2 і зверху вниз над травостоєм обмеженим квадратом, розглядають крізь комірки відомого масштабу, визначаючи відсоток покриття окремих видів, які є компонентами даного фітоценозу

|
Масштабні вилочки для обліку квадратів розміром 100x100 см 4%
від 1м2(А) і 50x50 см з площиною 10% від 0,25 м2 (Б) (за Раменським)
Крапчастий метод оцінки проективного покриття
Цей методзапропонував новозеландський учений Леві. Суть його полягає в наступному. В травостій фітоценозу втикають сталеві спиці, закріплені на планці завдовжки до 50 см з інтервалом 5 см,: підраховують кількість проколів стебел і листків або зіткненість їх зі спицями.
Підрахунок проколів при значних замірах і наступна статистична обробка масиву кількісних даних дає можливість судити про зімкнутість і проективне покриття фітоценозу.

Під поняттям "проективна рясність" розуміють проективне покриття окремих видів та груп рослин у фітоценозі. За великої кількості даних досягається висока точність визначення 0,1 – 0,05%, при побіжному підрахунку кількості видів: 0,3 – (0,5) – 1%. Проекційна рясність окремих видів або груп їх повинна в сумі дорівнювати проективній повноті, (тобто загальному і ярусному покриттю) угруповання.
Тому цей метод мало застосовують, а частіше користуються окомірним визначенням проективної рясності, яке потребує набагато менше часу. Щоправда, одержані дані будуть менш точними.
Горизонтальну проекцію крон дерев визначають окомірно, в частках від одиниці або у відсотках. За одиницю або 100% приймають таку зімкнутість крон, за якої просвіти між ними не перевищують 1/10 обстежуваної площі, а сума проекцій крон близько 9/10 її. Наприклад, якщо зімкнутість крон 0,5 це означає, що вона дорівнює половині повної зімкнутості деревостану.
Подібним способом визначають проекції крон окремих деревних порід за 10-бальною шкалою:
10 балів – повне покриття (100-90%),
9 –дуже велике покриття (90-80%),
8 – велике покриття (80-70%),
7 –досить велике покриття (70-60%)
6 –помірне покриття (60-50%),
5 – помітна зрідженість (50-40%),
4 – дуже велика зрідженість (40-30%),
3 – мале покриття (30-20%),
2 –дуже мале покриття (20-10%),
1 –майже відсутнє покриття (10%).
Сумарна проекція може бути більше 100%, якщо проекції одних особин перекривають проекції інших.
Проективне покриття в рослинних угрупованнях визначають для кожного ярусу. Так, у лісі проективне покриття окремо визначають для деревостану, кущів, травостою, мохово-лишайникового покриву; на болоті – для дерев, болотних кущів, травостою, сфагнових мохів.
Часткове проективне покриття створюють усі рослини одного виду. В багатовидових фітоценозах часто визначають часткове покриття, створеного групами окремих видів.
Видове проективне покриття – це часткове покриття, створене особинами одного виду.
Індивідуальне покриття – це проекція надземних частин однієї особини певного виду.
Справжнє, або дійсне, покриття – це покриття, утворене основами зрізаних стебел трав'янистих рослин і стовбурів деревних порід. Особливо чітко таке покриття виявляється в агрофітоценозах (наприклад, по стерні), після скошування травостою на луках, по пеньках після рубки деревостану.
Справжнє проективне покриття завжди буде менше загального чи ярусного проективного покриття, оскільки площа стовбурів менша, ніж площа проекцій крон.
Завдання.
1. Визначити загальне проективне покриття за допомогою сітки Л.Г. Раменського.
2. Визначити зімкнутість крон за допомогою зеркальної сітки.
3. Обрахувати відсоток покриття окремих видів у фітоценозі методом масштабної проекційної сіточки.
4. Визначити проективне покриття крапчатим методом.
5. Зробити висновки.
20
Лабораторна робота № 4
Тема: Застосування статистичних методів для
дослідження фітоценозів.
Мета: за допомогою статистичних методів визначити показники, що вказують на найбільш типовий рівень розвитку угруповання.
Обладнання: статистичні дані рослинних угруповань (таблиці), калькулятор.
Застосування статистичних методів в біології набуло таких особливостей, що можна вважати біологічну статистику невід'ємною частиною досліджень угруповань організмів.
В даній роботі використовуються статистичні методики П. Ф. Рокітцкого.
Рослинні організми характеризуються мінливістю та варіацією за різними ознаками. При вивченні біологічних угруповань, виявилось доцільним застосовувати статистичних дані.
Число одиниць сукупності називають об'ємом сукупності (n),
а варіацією – зміну значень ознаки. Варіанта (х і) - одне із значень, де і - означає порядковий номер варіанту.
Після підрахунку та розподілу всіх варіантів за класами утворюються ряди, в яких показано, як часто зустрічається певна варіанта. Класи з найбільшою частотою називаються модальні, а з найменшою – граничні.
Після підрахунку середнього арифметичного:

Необхідно вирахувати стандартне відхилення (варіанту):

та коефіцієнт варіації:

Вищезгадані характеристики дають можливість вичерпно охарактеризувати біологічні сукупності, які можуть бути представлені у вигляді ряду розповсюдження. Для цього ряду можна визначити статистичні показники, що вказують на найбільш типовий рівень розвитку угруповання, яке визначається, в сукупності ознаки і на всю ступінь варіації окремих одиниць сукупності навколо цього рівня. Але так як всі ці величини - це статистичні дані, то виникає питання наскільки вони достовірні.
Загальні закономірності випадкової варіації дають можливість підійти до питання про оцінку достовірності статистичних показників. Генеральна (х) та вибіркова (µ) сукупності характеризуються однаковими закономірностями випадкової варіації. Певні значення середніх арифметичних вибірок варіюють навколо середнього арифметичного генеральної сукупності. Варіація ж вибіркових середніх навколо μ може бути змінена своїм середнім квадратичним відхиленням, своєю σ. Ця сигма отримала назву середньої похибки, або середньої квадратичної помилки.
Інколи її ще називають стандартною похибкою (m) - вона вказує на ступінь зближення значень х та μ і вираховується за формулою:

З наведеної вище формули очевидно, що розмір визначеної середньої похибки залежить від σ і вибіркової популяції та її об'єму. Чим краще взята вибірка і чим більше її розмір, тим менша середня похибка. У зв'язку між статистичними показниками вибіркової та генеральної сукупностей віддзеркалюється, так званий, закон великих чисел – чим більше число n, деяких впадкових величин, тим їх середнє арифметичне ближче до середнього арифметичного генеральної сукупності, тим менше різниця між х та μ (Чебишев); при збільшенні n (n=∞), то х та μ прямують до 1, тобто до повної вірогідності. Для встановлення вірогідності різниці між середніми арифметичними використовують нормативне відхилення (t):
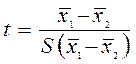
Де х1 – х2 – середнє арифметичне, S(х1– х2) – середнє відхилення. Якщо х1– х2= d, то


Якщо різниця перевищує свою похибку у 3 рази, то її вірогідність не менше 0,99, при n> 30, t = 2,58 (за таблицями).
При порівнянні двох груп з малими n помилка різниці вираховується за формулою:

якщо t ≥ 3, то різницю між сигмами можна вважати достовірно - ''суттєвою".
Завдання:
1. За допомогостатистичних даних рослинних угруповань (таблиці) визначити:
· середнє арифметичне значення одієї із ознак рослини (x);
· стандартне відхилення (варіанту) (σ);
· коефіцієнт варіації (V);
· стандартною похибкою (m);
· нормативне відхилення (t);
· помилку різниці (Sd).
2. Встановити найбільш типовий рівень розвитку угруповання.
3. Зробити висновки.
ЛАБОРАТОРНА РОБОТА № 5
Тема: визначення біомаси і продукції у рослинних угрупованнях
Мета: навчитись визначати біомасу і продукцію у рослинних угрупованнях.
Обладнання: описи рослинних угрупонь, база даних, калькулятор.
Кількісне співвідношення видів у фітоценозі визначають не лише шляхом підрахунку екземплярів того чи іншого виду але й за їхньою масою або продукцією. Для цього використовують різні способи: підрахунок та зважування у водоростей, зважування надземної частини у трав'янистих і дерев'янистих рослин
Ваговий метод обліку застосовується тоді, коли необхідно визначити рясність або кількісні параметри виду в господарько цінних або значимих фітоценозах, для визначення продуктивності природних кормових угідь, лісових насаджень, ягідників, обсяг заготівлі лікарської сировини тощо.
Визначають фітомасу як окремого виду, так і групи видів (наприклад, господарськи цінних рослин сінокосу чи пасовища), а нерідко й рослинного угруповання. Ваговий облік проводять, якщо треба визначити біомасу окремих ярусів чи функціональних біогеогоризонтів: крони, гілок, стовбурної деревини, урожай зерна, плодів, ягід.

Залежно від призначення (способу використання) біомасу зважують у сирому, повітряно-сухому і абсолютно сухому стані. У загальну масу органічної речовини включають масу наземної та підземної частини рослин. Для визначення маси підземної частини рослин роблять ґрунтові ями (розрізи) і на їхніх стінках за допомогою металічного шаблона або іншого пристрою вирізають ґрунтові моноліти на певній глибині (0–5см або 0–10см, 11–20см, 21–30см, 31–40см і глибше) або на глибині генетичних горизонтів. З вирізаних монолітів вибирають корені, промивають їх на ситах з певним діаметром отворів і висушують фільтрувальним папером. Нерідко при детальних дослідженнях їх розбирають за довжиною по фракціях: 0,1–0,5мм; 0,6–1,0мм; 1,1–2,0мм; 2,1–4,0мм; 4,1–6,0мм. Перші фракції складають поглинальні корінці (наприклад, у сосни (Pinus sylvestris), яка росте на болоті, вони зосереджені головним чином в 0–10см горизонті; до наступної фракції належать провідникові корені, точніше–корінці. Звичайно для відбору їх користуються мікроскопом або бінокулярною лупою.
Корені класифікують на живі, відмираючі та мертві для визначення функціональної активності кореневих систем у різних геогоризонтах.

Крім того, для всього угруповання або ж виду чи групи видів визначають загальну масу органічної речовини (живої і мертвої) в екосистемах.
Біомаса, або фітомаса – це загальна кількість органічної речовини, утвореної рослинами в екосистемі, за винятком мертвої речовини, яка синтезована за цей час рослинами, але не є частиною їх. Так, наприклад, відмираючі рослини болотних материнських фітоценозів в процесі їх мінералізації перетворюються в органічну масу – торф і не входять до складу фітомаси. Біомаса екосистем складається з біомаси популяцій видів, що створюють ці екосистеми. Тому за величиною біомаси того чи іншого виду можна до певної міри судити про його роль у фітоценозі та екосистемі.
Масу неживої органічної речовини утворює мертва органічна маса тваринного і рослинного походження – опад, відпад, трупи комах, червяківта інших організмів. Вона відокремлена від цілісного організму рослини та не належить до життєдіяльних органів.
Однією з характерних особливостей рослинного угруповання є здатність продукувати органічну речовину. В цьому процесі беруть участь всі рослинні й тваринні організми, що населяють фітоценоз. Рослинні організми шляхом фотосинтезу утворюють органічні речовини, первинну продукцію, на яку припадає до 80% загальної біомаси.
Вторинна продукція синтезується гетеротрофними організмами, ще менша кількість органічної речовини створюється хемотрофами.
А.Г. Воронов (1973) вважає, що для позначення конкретних кількісних показників доцільно вживати термін не "продуктивність", а "продукція" речовини. В геоботаніці розрізняють такі види продукції: загальна первинна, чиста первинна, або приріст, продукція опаду, продукція речовини.
Загальна первинна продукція, або брутто-продукція, звичайно розглядається, як сумарне збільшення фітомаси угруповання або популяції виду за одиницю часу (добу, вегетаційний період, рік тощо) на одиниці площі (м2, га), включаючи речовину, витрачену на дихання і процеси росту, а також кількість речовини опаду й екскретів гетеротрофних організмів.
Чиста первинна продукція, або нетто-продукція рослинного угруповання чи популяції виду – це фактичний приріст фітомаси за певний проміжок часу на одиниці площі, тобто кількість органічної речовини, акумульованої рослинами після часткового використання її на дихання.
Опад – це відмерла первинна продукція, тобто кількість органічної речовини, відмерлої в наземних частин рослин протягом певного часу на одиниці площі. Це відмерлі гілки, кора, суцвіття, опалі квітки, плоди тощо.
Відпад – частина опаду, яка складається з дерев і чагарників, відмерлих за певний час на одиниці площі. Виникає внаслідок зрідження деревостану, травостою.
Листковий відпад – це частина опаду, що складається з листків, приквітків, квіткових лусок, дрібних гілочок тощо, які відпали за певний проміжок часу на одиниці площі.
Коренепад – частина загального опаду, яку утворюють корені й підземні пагони, що відмерли у фітоценозі за певний час на одиниці площі.
Підстилка – наземний шар рослинних і тваринних решток різного ступеня мінералізації та потужності.
У геоботаніці прийняті ще такі поняття і терміни:
видова продукція – продукція створена одним якимось видом;
валова продукція – вся продукція;
корисна продукція – частина валової продукції, корисної в господарському відношенні;
валовий запас – загальний запас господарсько цінної біомаси (деревина в лісах, трав'яна маса на сінокосах тощо).
Наведені вище поняття, якими користуються для характеристики біомаси та фітомаси, дають можливість її оцінити як в цілому, так і окремі її компоненти, а також щодо виду, його участі у фітоценозі й фітоценозу взагалі. При цьому рослинні угруповання різняться за наземною та підземною фітомасою, співвідношенням господарськи цінних рослин.
Завдання.
1. Визначають фітомасу окремих видів у фітоценозі.
2. Визначити біомасу в сирому, повітряно-сухому і абсолютно сухому стані.
3. Визначити масу підземної частини рослин.
4. Вирахувати загальну масу органічної речовини, включаючи масу наземної та підземної частин рослин.
5. Дайте відповіді на запитання:
§ Що таке біомаса, або фітомаса?
§ Що таке загальна первинна продукція?
§ Що таке чиста первинна продукція?
§ Що таке опад?
§ Що таке відпад?
§ Що таке листковий відпад?
§ Що таке коренепад?
§ Що таке підстилка?
6. Зробити висновки.
ЛАБОРАТОРНА РОБОТА № 6
Тема: віковий спектр ценопопуляцій.
Мета: ознайомитись з віковим спектром ценопопуляцій.
Обладнання: описи рослинних угрупонь, база даних.
Кожна популяція представлена особинами, які в процесі онтогенезу в якийсь момент досягають певної фази (стадії) розвитку. Якщо в складі популяції є особини різного віку, для позначення їх правильніше буде скористатися терміном спектр вікових станів популяції, а не віковий склад. Виділяють такі вікові стани в життєвому циклі розвитку рослин: латентний, ювенільний, віргінільний, генеративний і сенільний (Злобін, 1989). Цю періодизацію з доповненням кожного періоду певними віковими станами можна представити у вигляді узагальненої таблиці:
Періодизація онтогенезу квіткових рослин
| Період | Віковий стан | Позначення |
| I. Латентний | 1. Насіння | L |
| II. Віргінільний | 2. Проростки 3. Ювенільні 4. Іматурні 5. Віргінільні (зрілі) | P J Im V |
| III. Генеративний | 6. Молоді генеративні 7. Середні генеративні 8. Старі генеративнi | G1 G2 G3 |
| IV. Сенільний | 9. Субсенільні 10. Сенільні 11. Відмираючі | SS S SC |
Якщо особини належать до одного абсолютного віку або одного вікового стану, їх називають віковою групою або когортою.
Якщо є точна інформація про календарний вік особин (наприклад, відомі строки посіву зернових або закладання розсадника чи посадки лісу) то доцільно вжити термін «віковий склад», а не «віковий спектр» популяції, оскільки в даному випадку всі особини мають однаковий вік і вся популяція є однаковою за віком.
Латентний період
Цей період охоплює період первинного стану спокою, коли особини представлені плодами, насінням, іншими рослинними зачатками (діаспорами). Тривалість цього періоду різна, так насіння одних видів: наприклад, осики (Populus), бука (Fagus), ялини (Рісеа) втрачає схожість за кілька днів, інших: наприклад, осок (Саrех), ситників (Juncus), костриць (Festuca) тощо зберігає її протягом тривалого часу.
Частина насіння (плодів, діаспор) дуже швидко втрачає схожість через несприятливі умови, частина гине або поїдається гризунами. Отже, не всі рослинні зачатки є життєздатними. Доведено, що основна маса їх зберігається в поверхневих шарах ґрунту та підстилці, а в нижніх – дуже мало.
Запас діаспор у ґрунті різниться за роками і за якісним складом, і за кількістю, цього запасу завжди вистачить для самовідтворення популяції та екосистем.
Насіннєва продуктивність фітоценозів та їхніх популяцій визначається багатьма факторами, такими, як історія фітоценозу, флористичний склад та кількісне співвідношення компонентів, умови місцезростання, можливості обнасінення, вплив тваринних організмів, занесення діаспор із суміжних територій, довговічність насіння, насіннєва продуктивність, господарська діяльність людини.
Окрему групу складають популяції однорічних рослин (польових, лучних, степових), насіння яких проростає лише за сприятливих умов. Так, насіння однорічних бобових, наприклад, горошка чотиринасінного (Vicia tetrasperma), г. волосистого (V. hirsuta), г. мохнатого (V. villosa) рясно проростає, якщо ґрунт має підвищену вологість.
У таких ценопопуляціях активний стан змінюється станом спокою: перший характеризується розвитком повноцінних зрілих особин, а другий – збереженням життєздатного насіння в ґрунті. Щоправда, можливі й відхилення від такого циклу. Наприклад, вовчки (Orobanche), які близько
150 років тому паразитували тільки на полинах, у наш час набули активного розвитку в культурі завдяки тому, що їхні ценолопуляції переходили до стану спокою при зміні полів у сівозмінах. Здатність насіння тривалий час зберігати схожість забезпечує високу стійкість видів у фітоценозах. Інакше кажучи, життєздатність насіння є чи не найголовнішою умовою (ознакою) стійкості ценопопуляції. Популяції, в яких усі види представлені лише насінням, тобто в період спокою перебуеають у латентному стані, називають латентними. В деяких видів чергування латентного й активного стану відбувається не щороку, а через певний період.
Віргінільний період
Віргінільний період досить довгий в онтогенезі деревних порід і багаторічних трав'янистих рослин. Він триває до першої стадії генеративного періоду й представлений ценопопуляціями, які знаходяться в стадії проростків, ювенільному та іматурному стані. Розглянемо окремо кожний віковий стан.
Стан проростків настає після завершення латентного періоду, коли особини популяції перебували в стані насінини. За сприятливих умов (відповідного температурного режиму, вологості та аерації ґрунту) насіння проростає, і починається якісно нова вікова стадія – стадія проростків. Т.А. Работнов (1978) до сходів відносить особини, котрі виникли в цьому вегетаційному сезоні з насіння або зачатків (бульбочок), що замінюють їх на початковому етапі життя. Такі особини морфологічно відрізняються від наступної вікової групи наявністю сім'ядолей, колеоптиле, колеоріози тощо. Оскільки проростки мають низьку адаптивну здатність до екстримальних умов навколишнього середовища, досить великий відсоток особин популяції гине на цій стадії. З розвитком кореневої системи і надземних частин кількість життєздатних сходів збільшується. Час перебування популяції односім’ядольних рослин у стані сходів, як правило, коротший, ніж у двосім’ядольних рослин, і людина враховує це при вирощуванні сільськогосподарських культур.
Ювенільна стадія настає в один і той же вегетаційний період, після завершення стадії сходів, тобто з віком особини переходять зі стадії сходів у якісно нову стадію – ювенільну. Ця стадія рослин є перехідною від сходів до сформованих віргінільних особин. В ювенільному стані рослини спочатку ще не мають повністю розвинених ані надземних органів – (листків, пагонів), ані підземних (коренів, кореневих систем), не створюють сформованих ценозів. Згодом вони вступають у фазу активного морфогенезу вегетативних органів, формуються адаптивні
адаптивні морфологічні й анатомічні ознаки, що забезпечують оптимальне їх функціонування. Ювенільна стадія – стан розвитку вегетативних органів особин, коли вони починають взаємодіяти міжсобою і в такий спосіб забезпечують формування окремих фаз фітоценозу на відкритих місцях або підросту в лісових та багаторічних лучних місцях. Особини багаторічних рослин розвиваються повільно і знаходяться в ювенільному стані від двох– трьох до п'яти–десятироків. Згодом вони швидко наростають і досягають стану зрілих віргінільних особин.

