




Артерии. Артерии не отличаются от артерий других локализаций и обладают средним мышечным слоем и адвентицией, содержащей коллагеновые и толстые эластические волокна (рис. 3.8.56). Мышечный слой от эндотелия отделен внутренней эластической мембраной. Волокна эластической мембраны переплетаются с волокнами базальной мембраны эндотелио-цитов [496, 959].
По мере уменьшения калибра артерии превращаются в артериолы. При этом исчезает сплошной мышечный слой стенки сосудов.
Сосуды и сосудистая оболочка глазного яблока


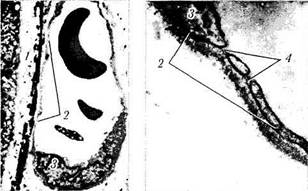
Рис. 3.8.56. Микроскопическое строение сосудов хорио-идеи:
а — артерия и вена хориоидеи крупного калибра (стенка артерии обладает толстым средним слоем и адвентицией); б, в — особенности ультраструктуры капиллярных сосудов хориоидеи (/—мембрана Бруха; 2 —эндотелиальная выстилка капилляра; 3 — ядро эндотелиальной клетки; 4 —дубликатура цитоплазмати-ческой мембраны с образованием «пор»)
Вены. Вены окружены периваскулярной оболочкой, вне которой располагается соединительная ткань. Просвет вен и венул выстлан эндотелием. Стенка содержит неравномерно распределенные гладкомышечные клетки в небольшом количестве. Диаметр самых больших вен равен 300 мкм, а самых маленьких, прека-пиллярных венул, — 10 мкм [154, 1028].
Капилляры. Капилляры хориокапиллярного слоя сосудистой оболочки имеют довольно большой просвет, позволяющий проходить нескольким эритроцитам. Выстланы они эндоте-лиальными клетками, снаружи которых лежат перициты (рис. 3.8.56, б, в). Количество перицитов на одну эндотелиальную клетку хориокапиллярного слоя довольно велико. Так, если в капиллярах сетчатки это соотношение равно 1:2, то в сосудистой оболочке— 1:6 [370, 708, 933]. Перицитов больше в фовеолярной области. Перициты относятся к сократительным клеткам и участвуют в регуляции кровоснабжения. Особенностью капилляров хориоидеи является то, что они фенестрированы, в результа-
те чего их стенка проходима для маленьких молекул, включая флюоросцеин и некоторые белки [ill, 1007]. Диаметр пор колеблется от 60 до 80 мкм. Закрыты они тонким слоем цитоплазмы, утолщенной в центральных участках (30 мкм). Фенестры располагаются в хориока-пиллярах со стороны, обращенной к мембране Бруха [496, 527] (рис. 3.8.57, в). Между эндо-телиальными клетками артериол выявляются типичные зоны замыкания.
Межклеточные контакты эндотелиальных клеток хориокапилляров особого типа. Близкие по строению контакты выявляются в синусоидах печени и венулах брыжейки [1208]. Выявляются неравномерно распределенные зоны замыкания и десмосомы, которые не полностью герметичны [875, 1028]. Между эндотелиальными клетками и перицитами существуют щелевые контакты [1028].
Иннервация сосудистой оболочки. Сосудистая оболочка иннервируется симпатическими и парасимпатическими волокнами, исходящими из ресничного, тройничного, крылонебного и верхнего шейного ганглиев (рис. 3.8.57). В глазное яблоко поступают они с ресничными нервами.

Рис. 3.8.57. Особенности вегетативной иннервации уве-ального тракта глаза:
/ — крылонебный ганглий; 2 —верхний шейный симпатический
ганглий; 3 — ресничный ганглий; 4 — тройничный ганглий; 5 —
пятый нерв
В строме сосудистой оболочки каждый нервный ствол содержит 50—100 аксонов, теряющих миелиновую оболочку при проникновении в нее, но сохраняющих шванновскую оболочку.
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 Постганглионарные волокна, исходящие из ресничного ганглия, остаются миелинизиро-ванными.
Постганглионарные волокна, исходящие из ресничного ганглия, остаются миелинизиро-ванными.
Сосуды надсосудистой пластинки и стромы сосудистой оболочки исключительно обильно снабжены как парасимпатическими, так и симпатическими нервными волокнами (рис. 3.8.58).
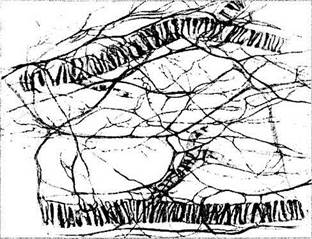
Рис. 3.8.58. Особенности распределения нервных волокон между сосудами сосудистой оболочки
Симпатические адренергические волокна, исходящие из шейных симпатических узлов, обладают сосудосуживающим действием.
Парасимпатическая иннервация сосудистой оболочки исходит от лицевого нерва (волокна, идущие из крылонебного ганглия), а также из глазодвигательного нерва (волокна, идущие из ресничного ганглия).
Последние исследования значительно расширили наши знания относительно особенностей иннервации сосудистой оболочки. У различных животных (крыса, кролик) и у человека артерии и артериолы сосудистой оболочки содержат большое количество нитрэргических и пептидэргических волокон, образующих густую сеть. Эти волокна приходят с лицевым нервом и проходят через крылонебный ганглий и не-миелинизированные парасимпатические ветви от ретроглазного сплетения [328, 1202]. У человека, кроме того, в строме сосудистой оболочки имеется особая сеть нитрэргических ган-глиозных клеток (положительны при выявлении НАДФ-диафоразы и нитроксидной синте-тазы), чьи нейроны связаны друг с другом и с периваскулярной сетью (рис. 3.8.59). Отмечено, что подобное сплетение определяется только у животных, имеющих фовеолу.
Ганглиозные клетки сконцентрированы в основном в височных и центральных областях сосудистой оболочки, по соседству с макуляр-ной областью. Общее количество ганглиозных клеток в сосудистой оболочке порядка 2000. Распределены они неравномерно. Наибольшее их количество обнаруживается с темпоральной
стороны и центрально. Клетки маленького диаметра (< 10 мкм) располагаются по периферии [328]. Диаметр ганглиозных клеток увеличивается с возрастом, возможно, из-за накопления в них липофусциновых гранул.
В нейронах выявлены нитрэргические трансмиттеры. Подобные нейротрансмиттеры обеспечивают расширение сосудов. Обнаруживаются они в периваскулярных нервах различных органов [116, 757, 787, 1081]. Этот медиатор вызывает также расслабление гладких мышц различных органов, например кишечника и трахеи [423], желчного пузыря [1059].
В некоторых органах типа сосудистой оболочки нитрэргические нейротрансмиттеры выявляются одновременно с пептидэргическими, также обладающими сосудорасширяющим действием [365, 614, 739, 1059]. Пептидэргичес-кие волокна [1118], вероятно, исходят из крылонебного ганглия и проходят в лицевом и большом каменистом нерве [1118]. Вероятно, что нитро- и пептидэргические нейротрансмиттеры обеспечивают вазодилятацию при стимуляции лицевого нерва.
Периваскулярное ганглиозное нервное сплетение расширяет сосуды сосудистой оболочки, возможно регулируя кровоток при изменении внутриартериального кровяного давления. Оно защищает сетчатку от повреждения тепловой энергией, выделяющейся при ее освещении. Flugel et al. [328] предложили, что ганглиозные клетки, расположенные у фовеолы, защищают от повреждающего действия света именно тот участок, где происходит наибольшая фокусировка света. Выявлено, что при освещении глаза существенно увеличивается кровоток в прилежащих к фовеоле участках сосудистой оболочки.

Рис. 3.8.59. Ганглиозная клетка сосудистой оболочки типичного строения, к которой подходит и контактирует нервное волокно:
/ — ганглиозная клетка; 2 —крупное ядрышко ганглиозной клетки; 3 — нервное волокно
Сосуды, и сосудистая оболочка глазного яблока
 Особенности кровообращения в сосудистой оболочке. Особенности кровообращения уве-ального тракта изучались интенсивно на протяжении многих лет как в эксперименте, так и в клинике. В 1975 г. Bill [114] суммировал имеющиеся данные и привел свою концепцию физиологии хориоидеи.
Особенности кровообращения в сосудистой оболочке. Особенности кровообращения уве-ального тракта изучались интенсивно на протяжении многих лет как в эксперименте, так и в клинике. В 1975 г. Bill [114] суммировал имеющиеся данные и привел свою концепцию физиологии хориоидеи.
У обезьян хориоидальный кровоток исключительно интенсивный, приблизительно в 20 раз выше, чем в сосудах сетчатой оболочки (радужка — 8 ± 1 мг/мл; ресничное тело — 81 ±6 мг/мл; сосудистая оболочка — 677 ± ±67 мг/мл; сетчатка — 34 ± 2 мг/мл). Поскольку интенсивность артериального кровотока столь высока, насыщенность кислородом венозной крови только на 3% ниже, чем насыщенность артериальной крови. И это несмотря на то, что кислород отдается наружной части сетчатой оболочки. Артериовенозные анастомозы играют небольшую роль в поддержании высокой насыщенности кислородом венозной крови. Предлагается, что высокий уровень увеаль-ного кровотока обеспечивает терморегуляцию внутриглазных оболочек, компенсируя снижение температуры в переднем отделе глаза и предотвращая перегревание сетчатки при ее освещении светом.
Регуляция кровотока. Механизмы регуляции кровотока в сетчатке и сосудистой оболочке существенно отличаются. Если в сетчатке преобладают механизмы ауторегуляции, то в хориоидее эти функции берут на себя симпатические нервные сплетения.
Интенсивность кровотока в сетчатке незначительно увеличивается при повышении концентрации рСО2 [347], а гипероксия вызывает небольшое сужение сосудов. При этом интенсивность кровотока снижается. Особенностью кровообращения сетчатки является и то, что на него не влияет изменение внутрисосудистого давления, что наблюдается, например, при изменении внутриглазного давления.
Кровообращение в хориоидее также усиливается при увеличении концентрации рСО2, но более значительно. При повышении парциального давления кислорода интенсивность кровотока практически не изменяется [112, 113].
Кровообращение хориоидеи не автономно, а регулируется нервными механизмами [248]. Ауторегуляция кровообращения выявлена только в сосудах ресничного тела и радужки. При стимуляции симпатической нервной системы наступает уменьшение просвета сосудов хориоидеи. При этом падает внутриглазное давление из-за уменьшения объема крови. Подобная реакция характерна для а-адренэргического типа иннервации [112, 113].
Сосуды хориоидеи находятся обычно в состоянии небольшого сокращения (сосудосуживающий тонус). Предполагают, что такое состояние защищает сетчатку от гиперперфузии сосудов, наблюдающейся при ряде заболеваний,
сопровождающихся повышением внутриартери-ального давления [119]. Вазомоторные терминалы заканчиваются в основном на артериолах и реже на артериях. Иннервируются также вены и венулы. Отсутствует иннервация хорио-капилляров [933]. На холинэргическую стимуляцию сосуды хориоидеи отвечают расширением просвета [1037].
В увеальном тракте выявлены также нит-ро- и пептидэргические волокна, обладающие сосудорасширяющим действием [933]. Подходят они к глазу по ходу лицевого нерва, образуя синапсы в крылонебном ганглии [328, 786, 1037, 1118].
Капилляры сосудистой оболочки и ресничных отростков напоминают таковые слизистой оболочки кишечника и почки. Исследование проницаемости этих капилляров выявило, что стенка пропускает большие молекулы. Дальнейшее продвижение молекул из сосудистой оболочки в сетчатку невозможно в результате наличия между пигментными клетками эпителия сетчатки плотных межклеточных контактов [222, 399, 1016]. Утечка белка из просвета капилляров сосудистой оболочки или ресничных отростков превышает подобную утечку в почках в пять раз, а в сердечной и скелетной мышцах в десять [120]. Благодаря такой высокой пропускной способности стенки сосудов, концентрация IgG в строме ресничных отростков и строме сосудистой оболочки составляет 60— 70% концентрации этого белка в плазме крови. Это свойство создает высокое осмотическое давление в ткани сосудистой оболочки (превышает давление сетчатки примерно на 15 мм ртутного столба). Разница в осмотическом давлении между сосудистой оболочкой и сетчаткой вызывает фильтрацию жидкости из сетчатки по направлению к сосудистой оболочке и является силой, которая придавливает сенсорную часть сетчатки к пигментному эпителию.
Наличие высокой проницаемости сосудов хориоидеи способствует транспорту в сетчатую оболочку витамина А, находящегося в макромо-лекулярном комплексе ретинол-связанного белка с преальбумином. Возможность выхода такой большой молекулы обеспечивается наличием фенестр. Высока пропускная способность сосудов и для низкомолекулярных веществ типа глюкозы. Причем она более чем в двадцать раз выше относительно сосудов мышцы сердца и в восемьдесят относительно сосудов скелетной мышцы. Это резко отличает сосуды хориоидеи от сосудов сетчатой оболочки.
Ишемия хориоидеи. На протяжении многих десятилетий непонятной оставалась причина развития ишемии хориоидеи при столь высокой насыщенности ее анастомозирующими сосудами. Причем участки ишемии хориоидеи строго очерчены [1198, 1199].
Механизмы развития локальной ишемии были непонятны и по следующим причинам:

