




В угрупуваннях існує міжвидова конкуренція, тобто боротьба за ресурси всередині угрупування.
Існують наступні типи взаємодії двох популяцій в угрупуваннях:
- НЕЙТРАЛІЗМ, при якому асоціація двох популяцій не відбивається на жодній з них.
- Взаємне КОНКУРЕНТНЕ придушення, при якому обидві популяції активно придушують одна одну.
- Конкуренція за ЗАГАЛЬНИЙ РЕСУРС, при якій кожна популяція посередньо негативно впливає на іншу в боротьбі за функціональний ресурс. (Більшість видів існує саме так, а не як 2).
- АМЕНСАЛІЗМ, при якому одна популяція придушує іншу, але сама не зазнає заперечного впливу.
- ПАРАЗИТИЗМ (особини – паразити менші за особин хазяїна) і хижацтво (особини – хижаки звичайно більше особин – жертв), при яких одна популяція несприятливо впливає на іншу, нападаючи безпосередньо на неї, але, тим не менше, сама залежить від об'єкта свого нападу.
- КОМЕНСАЛІЗМ, при якому одна популяція витягає пожиток з об'єднання, а для іншої це об'єднання байдуже.
- ПРОТОКООПЕРАЦІЯ, при якій обидві популяції одержують від асоціації пожиток, але ці стосунки не обов'язкові.
- МУТУАЛІЗМ, при якому зв'язок популяцій сприятливий для зростання і виживання обох, причому в природних умовах не одна з них не може існувати без іншої.
Із зазначених типів взаємодій популяцій найбільший практичний інтерес представляють конкуренція і хижацтво.
Для з'ясування дії різноманітних факторів на зміну кількості одного виду використовують математичні моделі у формі диференціальних рівнянь швидкостей зростання популяції, на яку впливає інша популяція.
dN1/dt = r1*N1 – r1/K1 – N12 – a*N1*N2, (3.3)
де r – репродуктивний потенціал;
К – стабільна кількість, до якої прямує популяція у даних умовах (ємність середовища);
N1, N2 – чисельність популяцій 1 і 2;
a – коефіцієнт, що визначає напруженість межвидової конкуренції;
dN1/dt – швидкість зростання популяції 1;
r1*N1 - максимальна швидкість зростання популяції 1;
r1/K1 – визначає напруженість внутрішньовидової конкуренції.
З рішень цих рівнянь випливає, що швидкість зростання кожної популяції дорівнює максимальній швидкості зростання мінус вплив власної кількості (напруженість внутрішньовидової конкуренції) і мінус вплив іншого виду (напруженість міжвидової конкуренції). Причому останній параметр (а) може бути позитивним (див. типи взаємодій двох популяцій в угрупуваннях п.п. 2, 3, 4*, 5), від'ємним (п.п. 6*, 7, 8,) і рівним нулю (1, 4*, 6*), (де * для одної з двох взаємодіючих популяцій).
При розгляді взаємодії великого числа популяцій рівняння складають відповідну систему рівнянь.
Якщо обмежитися взаємодією двох видів, можна записати друге рівняння:
dN2/dt = r2*N2 – r2*N2/K2 – b*N2*N1, (3.4)
де b – коефіцієнт конкуренції, що визначає вплив виду 1 на вид 2.
Система рівнянь для двох популяцій (3.3) – (3.4) носить назву рівнянь Лоткі – Вольтерра, отриманих ними незалежно один від одного у 1925 і 1926 р.р. і використовуваних В.Вольтерра для пояснення коливань рибних уловів в Адріатичному морі. Відтоді ці рівняння були предметом численних досліджень.
Один з основних результатів полягає в тому, що при конкуренції і відсутності сховищ, один з видів з необхідністю вимирає, а інший досягає стійкого стану. І це відповідає як експериментальним даним, так і реальному розвитку природних екосистем.
В системі "хижак – жертва" картина цілком інша. За відсутності внутрішньовидової конкуренції (К1 = К2 = ¥) і конкурентного впливу жертви на хижака (b = 0), але наявності залежності народжуваності хижака (В2) від кількості жертви (В2 = Kn*a*N1, де Кn – коефіцієнт корисної дії переробки біомаси жертви на біомасу хижака) одержуємо класичну модель В.Вольтерра
dN1/dt = r1*N1 – a*N1*N2
dN2/dt = (Kn*a*N1 – D2)*N2, (3.5)
де B,D – функції народжуваності та смерті;
a*N1 – швидкість споживання жертви одним хижаком.
Аналіз цієї моделі дозволяє зробити висновок, що в системі "хижак – жертва" за відповідних умов (див. вище) можливий стійкий режим коливань кількості жертв N1 і хижаків N2.
Суцільна крива на рис. 3.4. має назву граничний стійкий цикл. Вказано напрямок його проходження (стрілки) і розмітка по часу, наприклад, в роках (цифри). Швидкість проходження циклу по даній дільниці визначається похідною до кривої на цій же дільниці. Згідно із стійким циклом відбуваються взаємопов'язані коливання кількості популяцій жертви і хижака з одним періодом (в даному випадку 12 років). Такого роду залежності були реально виявлені. Вони відповідають найважливішій властивості живої природи – самоорганізації. Справді, цей цикл називається граничним, тому що до нього будуть прямувати всі інші траєкторії співвідношень кількості хижаків і жертв (пунктирні лінії), незалежно від їх початкового співвідношення. Таким чином, хижак виявляє якби стабілізуючий вплив на кількість жертви, не допускає її вибухового збільшення і загибелі надалі. Тобто, вовки і лисиці теж потрібні для стійкого існування зайців і козуль.
Дані розв'язки є детерміністськими, а реальне середовище носить багато в чому випадковий характер. В природі смертність і, особливо, народжуваність підлягають випадковим збуренням. Причому випадкові збурення можуть бути значними по амплітуді і протидіючими на два взаємодіючих видів. В цьому випадку за несприятливих обставин, тобто малої кількості одного з видів, може статися його вимирання. Такого роду ситуації неодноразово виникали і, нажаль, виникають в природі. Таким чином, чим менше кількість видів, більше розмах протилежних випадкових збурень кожного з видів, тим скоріше станеться вимирання одного з них, руйнування угрупування.
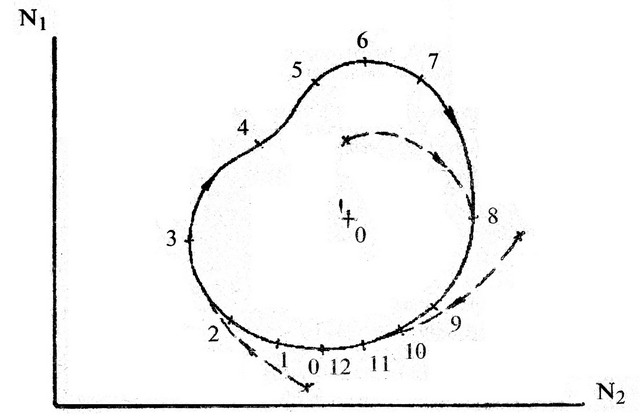
Рис. 3.4. Залежність кількості популяцій в системі "хижак – жертва".
Розрахунки на моделях, що враховують випадкові збурення, показали, що угрупування, що складається з хижака і жертви, не може стабілізуватися у випадковому середовищі при низькій смертності хижака поряд з високою доступністю жертви чи при високій смертності хижака і важкодоступності жертви. Такий висновок інтуїтивно зрозумілий і відповідає реальним ситуаціям в природі.
Треба також визначити, що використання математичних моделей дозволило сформулювати, кількісно визначити і оцінити вплив на популяції і угрупування двох найважливіших біотичних факторів екологічного середовища – внутрішньовидової і міжвидової конкуренції.
Людина, на жаль, ще дуже повільно навчається бути "ощадливим хижаком", тобто, не знищувати свою жертву повністю при експлуатації її, як ресурсу. Теоретично оптимальне вилучення з експлуатованої популяції (наприклад, риби з озера) повинно діятися до рівня, при якому подальша швидкість зростання популяції максимальна. Реально оптимальний вихід живої продукції забезпечується в обсязі кілька меншому половини граничної кількості популяції при даних умовах середовища пробування.
Взагалі, в екології властивість будь-якої системи відновлюватися після того, як її структура і функції були порушені, має назву пружня стійкість біосистеми. Це поняття тісне пов'язано із властивістю живої матерії до самоорганізації (рис. 3.4). Спроможність же біосистеми чинити опір, підтримуючи свою структуру і функції незмінними називають резистентною стійкістю. Одна і таж біосистема володіє, як правило, тільки одним типом підтримання стабільності.
Ще одне виключно важливе поняття в біоценології – екологічна ніша. Вона включає в себе не тільки фізичний простір, але і певну частину екологічного середовища, що описується, як відомо абіотичними і біотичними факторами. Отже екологічна ніша даного виду визначається не тільки місцем, де він живе, але і його вимогами до навколишнього середовища (температура, вологість і т.д.), а також взаємовідносинами з іншими видами при здобуванні їжі. Конкуренція за один і той же харчовий ресурс призводить до зменшення ніші одного з видів чи навіть його повному витисненню з даної ніші. Витиснення одного виду іншим може статися і в наслідок конкуренції по будь-якому параметру екологічної ніші, наприклад, кількості сонячної енергії, що надходить (у рослин). Зона екологічної толерантності виду відповідає ширині екологічної ніші по даному фактору. Для характеристики ніші використовують ще її перекривання нішами сусідніх видів, без чого не обходяться реальні біогеоценози. Види, що займають однакові ніші в різних географічних краях, мають назву екологічні еквіваленти.
Визначая біогеоценоз, як сукупність всіх видів, що населяють певну територію, ми фактично не наклали ніяких обмежень на географічні характеристики цієї території. Разом з тим, межі природних біосистем не можуть бути задані довільно. Тому упроваджено поняття, щоб уточнити раніше використовуваний нами термін. Біогеоценозом називається сукупність популяцій всіх видів живих організмів, які населяють певну географічну територію, що відрізняється від сусідніх територій за хімічним складом грунтів, вод, а також за рядом фізичних показників (висота над рівнем моря, величина сонячного опромінення і т.д.). Тобто, простір, що займає біогеоценоз, відмежований істотною зміною на його межі хоча б одного фактора екологічного середовища.
Різноманітність організмів на межі біогеоценозів має тенденцію до збільшення, що відомо під назвою крайового ефекту. Типовим прикладом може служити узлісся, де різноманітність рослин значно більше, ніж в лісі і степу.
Біогеоценоз і його середовище пробування разом виставляють собою біосистему самого високого рівня організації (рис. 3.1), що має назву екологічна система чи екосистема. Поняття часто використовується і без обмежень на характеристики простору і кількості видів, які його населяють, тобто може розглядатися довільний географічний простір з довільним числом видів.
Екосистема в більш точному розумінні (що має у вигляді біотичного компоненту біогеоценоз) володіє цікавою властивістю, що має назву екологічна сукцесія, яка відбувається в зміні по часу її видової структури. У відсутності зовнішніх порушуючих процесів сукцесія уявляє собою спрямований передбачений процес. Більш чи менш неперервна зміна видів у часі нагадує естафету. Внаслідок з'являються послідовності угрупувань, що називаються серіями, які також замініють один одного в даному географічному просторі. Для останньої серії в цьому ланцюзі розвитку екосистеми використовують термін – клімакс. Він теоретично повинен бути постійним в часі і існувати до того часу, поки його не порушать сильні зовнішні збурення.
Сукцесія, що починається на ділянці, яка раніше не була зайнята (потік застиглої лави), має назву первинна сукцесія. Сукцесія, що починається на площі.з якої усунено колишнє суспільство (лісне вирубування), має назву вторинна сукцесія.
На ранніх стадіях сукцесії переважає r – відбір, тобто успішно уселяються види з високою швидкістю розмноження і зросту. Напроти, для пізніх стадій сукцесії характерний k – відбір, тобто переважають види з низьким потенціалом зростання, але з більш високою здібністю до виживає мості в умовах конкуренції. Типовий ряд вторинної сукцесії в передгірному краї включає стадії: голе поле, степ, злаки і чагарники, сосновий ліс і, через 150 років, дубовий клімаксний ліс.

