




1. Кэпирование (англ. cap - шапка) – происходит еще во время транскрипции, состоит в том, что к 5’‑трифосфату концевого нуклеотида пре‑мРНК присоединяется 5’‑углерод N7‑метил-гуанозина. «Кэп» необходим для защиты молекулы РНК от 5’‑3’‑экзонуклеаз.
2. При транскрипции зон ДНК, несущих информацию о белках, образуются гетерогенные ядерные РНК, по размеру намного превосходящие мРНК. Дело в том, что из-за мозаичной структуры генов эти гетерогенные РНК включают в себя информативные (экзоны) и неинформативные (интроны) участки. При особом процессе – сплайсинге (англ. splice – склеивать встык) происходит удаление интронов и сохранение экзонов. 
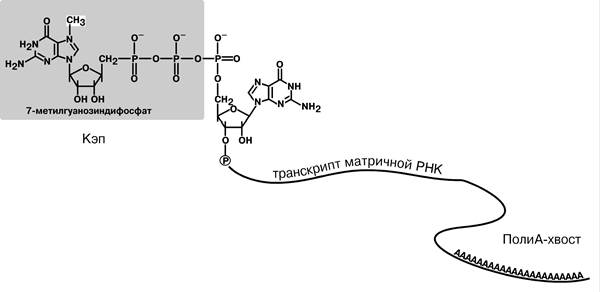 |
3. Полиаденилирование – при помощи полиаденилат-полимеразы с использованием молекул АТФ происходит присоединение к 3’‑концу 100‑200 адениловых нуклеотидов, формирующих поли (А)‑хвост.
Процессинг предшественника рРНК
Предшественники рРНК являются более крупными молекулами по сравнению со зрелыми рРНК
У прокариот большая прерибосомная 30S‑РНК расщепляется специфичными нуклеазами с образованием 5S‑рРНК, 16S‑рРНК, и 23S‑рРНК.
У эукариот большая прерибосомная 45S‑РНК расщепляется специфичными нуклеазами с образованием 5,8S‑рРНК, 18S‑рРНК, и 28S‑рРНК.
Процессинг предшественника тРНК
1. Формирование на 3’‑конце последовательности Ц‑Ц‑А. Для этого у одних пре‑тРНК с 3’-конца удаляются лишние нуклеотиды до «обнажения» триплета Ц‑Ц‑А, у других идет присоединение этой последовательности.
2. Формирование антикодоновой петли происходит путем сплайсинга и удаления интрона в средней части пре‑тРНК.
3. Модификация нуклеотидов путем дезаминирования, метилирования, восстановления. Например, образование псевдоуридина и дигидроуридина.
Регуляция транскрипции
Прокариоты
Регуляция биосинтеза белка у прокариот осуществляется на уровне транскрипции мРНК. В настоящее время принята теория оперона, сформулированная Франсуа Жакобом и Жако Моно. В основе теории лежат следующие понятия:
· конституитивные ферменты – те, которые присутствуют в клетках всегда, независимо от ее активности
· индуцибельные ферменты – те, которые синтезируются при появлении субстрата
· оперон – группа тесно связанных между собой генов (несколько структурных генов и один ген-оператор), которые регулируют образование ферментов в организме.
·
 |
ген-регулятор – ген, регулирующий работу оперона, не входящий в его состав.
Лактозный оперон
При изучении E.coli было замечено, что активность одного из ферментов катаболизма лактозы низка, если в среде имеется глюкоза. При отсутствии же глюкозы и при наличии лактозы активность фермента резко повышается. На основании этих наблюдений была предложена схема регуляции оперона по механизму индукции.
В отсутствие лактозы активный белок-репрессор связывается с оператором и блокирует синтез мРНК, кодирующей ферменты катаболизма лактозы. В результате эти ферменты не образуются.
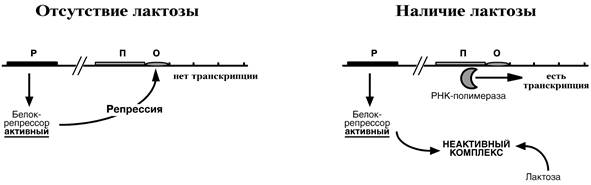 |
Если глюкозы нет, и есть лактоза, то последняя связывается с белком-репрессором и модифицирует его, не позволяя связаться с геном-оператором. Это позволяет РНК‑полимеразе считывать информацию, отвечающую за синтез ферментов катаболизма лактозы, и синтезировать мРНК.
Таким образом, лактоза является индуктором транскрипции.
Триптофановый оперон
Функционирование триптофанового оперона, в некотором смысле, противоположно работе лактозного. В данном случае, в отличие от лактозного оперона, белок-репрессор синтезируется в неактивном состоянии и не может заблокировать транскрипцию генов, кодирующих ферменты синтеза триптофана. Синтез аминокислоты будет в клетке продолжаться до тех пор, пока в среде не появится триптофан.
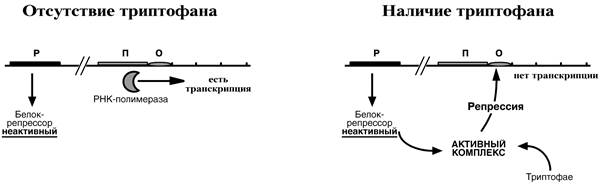 |
Триптофан соединяется с белком-репрессором и активирует его. Далее такой активный комплекс присоединяется к гену-оператору и блокирует транскрипцию. Таким образом, при наличии триптофана в среде, прекращается его внутриклеточный синтез, экономятся ресурсы и энергия бактериальной клетки.
В этом случае триптофан является репрессором транскрипции. Регуляция осуществляется по механизму репрессии.
Эукариоты
Существенное усложнение эукариотических организмов повлекло за собой появление новых способов регуляции активности матричных биосинтезов:
Амплификация – это увеличение количества генов, точнее многократное копирование одного гена. Естественно, все полученные копии равнозначны и одинаково активно обеспечивают транскрипцию.
Например, противоопухолевый препарат метотрексат препятствует работе дигидрофолатредуктазы, фермента, необходимого для синтеза дезоксирибонуклеотидов. При этом в опухолевых клетках происходит амплификация гена этого фермента, что приводит к многократному увеличению синтеза дигидрофолатредуктазы и невосприимчивости опухолевых клеток к метотрексату.
Энхансеры (англ. to enhance - усиливать) – участки ДНК в 10‑20 пар оснований, способные значительно усиливать экспрессию генов той же ДНК. В отличие от промоторов они значительно удалены от транскрипционного участка и могут располагаться от него в любом направлении (к 5’‑концу или к 3’‑концу). Сами энхансеры не кодируют какие-либо белки, но способны связываться с регуляторными белками.
Сайленсеры (англ. silence - молчание) – участки ДНК, в принципе схожие с энхансерами, но они способны замедлять транскрипцию генов, связываясь с регуляторными белками.
Перестройка генов. Нуклеотидные последовательности, кодирующие белковую молекулу могут оказаться разделенными на отдельные сегменты, не связанные между собой. Например, иммуноглобулины состоят из тяжелой и легкой цепей, каждая из которых включает собственные вариабельную и константную части. Существует множество вариантов как вариабельной, так и константной частей. Генетическая информация об этих вариантах локализована подчас в разных хромосомах. При дифференцировке В‑лимфоцитов значительно удаленные сегменты генетического материала переносятся и группируются рядом – происходит генетическая рекомбинация.
Процессинг мРНК – некоторые пре‑мРНК подвергаются разным вариантам сплайсинга (альтернативный сплайсинг) в результате чего образуются разные мРНК, и соответственно, белки с разной функцией. Примером может служить образование двух типов тяжелых цепей IgM в В‑лимфоцитах, один из которых удерживает IgM на мембране, другой позволяет антителу нормально секретироваться наружу.
Изменение стабильности мРНК – чем выше продолжительность жизни мРНК в цитозоле клетки, тем больше соответствующего белка наработается. Например, установлено, что при наличии пролактина в клетках молочной железы время полужизни мРНК белка казеина значительно увеличивается, а эстрадиол продлевает время полужизни мРНК белка вителлогенина в десятки раз.
Транспорт из ядра в цитоплазму –

