




Елементи промотора
Геном дріжджів включає близько 5000 генів, що кодують білки. Вони щільно розміщені на 16-ти лінійних хромосомах. Дріжджові гени всередньому транскрибуються 5-10 разів протягом кожного клітинного циклу, результатом чого є наявність стійкого пулу мРНК у кількості 1-2 молекули на клітину. Кількості утворених мРНК різних генів можуть відрізнятись більше, ніж на 2-3 порядки у клітинах. Деякі гени транскрибуються зі зміненою інтенсивністю в залежності від певних фізіологічних умов, а групи генів можуть піддаватись координованій регуляції [2]. Незважаючи на ці транскрибційні особливості, майже всі дріжджові промотори складаються з трьох основних елементів:
1. UAS – upstream елементи;
2. ТАТА – блок;
3. I – ініціатор [7].
Upstream елементи – це короткі (10-30 п.н.) промоторні специфічні послідовності ДНК, що розміщені 50-100 п.н. upstream від сайту ініціації. UAS дріжджів аналогічні, і в багатьох випадках гомологічні енхансерам ссавців. Однак, хоч вони й функціонують в обидвох напрямах і на різній віддалі upstream від сайту ініціації транскрипції, проте UAS не здатні активувати транскрипцію в положенні downstream. Upstream елементи розпізнаються ДНК-зв’язуючими білками, що визначають частково регуляторні особливості даних промоторів. Цікаво, що близько 20% промоторів дріжджів містять гомополімерні dA: dT треки, що функціонують як UAS завдяки своїй структурі ДНК, без взаємодії зі специфічними ДНК-зв’язуючими білками.
Більшість промоторів дріжджів містить ТАТА-блоки, консенсус яких ТАТААА. Вони важливі для ініціації транскрипції. Цей елемент розміщений 40-120 п.н. upstream від сайту ініціації, точна віддаль не відіграє важливої ролі у геномі дріжджів, на відміну від геному ссавців. ТАТА-елементи розпізнаються ТАТА-зв’язуючими білками (ТBP (TATA binding proteins), що є компонетами РНК-полімерази II та необхідні для транскрипції [7]. Промотори, що втратили ТАТА-елементи теж містять ТBP, але в цьому випадку дані білки зв’язуються з ДНК-послідовністю неспецифічно і стаблізуються завдяки білок-білковим взаємодіям з іншими транскрипційними факторами.
Ініціаторні елементи у дріжджів визначають місце початку транскрипції. Вони, на відміну від upstream і ТАТА-елементів, не мають певної консенсусної послідовності. Вплив ініціаторних елементів на рівень транскрипції є незначним, оскільки вони лише визначають, де вона розпочнеться [5, 7].
Деякі дріжджові промотори містять ще й елементи негативної регуляції – оператори, що репресують транскрипцію. Здатністю проявляти свій вплив незалежно від напряму на різних віддалях upstream від ТАТА-елементів, оператори нагадують вищезгадані UAS. Деякі оператори здатні репресувати транскрипцію навіть у положенні upstream від UAS, реперсія є значно ефективнішою, якщо оператор знаходиться між UAS і ТАТА-елементом [5].
У S. cerevisiae виявлено ген металотіонеїну, індукція транскрипції якого лежить в основі захисту клітин від токсичного впливу важких металів. Він не є життєво необхідним, але захищає дріжджі від токсичної дії купруму [3]. Дослідження промотора дозволило ідентифікувати два елементи, чутливих до міді, що знаходяться між –200 і –100 парами нуклеотидів, і звичайний ТАТА-блок поблизу –30 пари, відраховуючи з точки початку транскрипції (рис. 1).
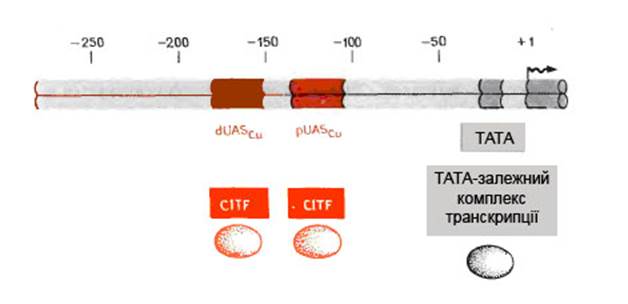
Рис. 1. Організація регуляторних елементів промотора гену
металотіонеїну S. cerevisiae
Ці два елементи, що активуються купрумом (UASCu), розміщені перед промотором і представлені недосконалими повторами, довжиною від 32 до 34 п.н. Це сайти зв’язування фактора купрум-індукованої транскрипції (CITF). Елемент ТАТА детермінує збирання комплексу транскрипції. Фактор CITF, ген якого (ACE1) влалось клонувати, секвенувати та експресувати, приєднується до UASCu лише в комплексі з міддю. Домени зв’язування міді та ДНК у CITF локалізовані на N-кінці молекули білка. Залежність зв’язування білку з UASCu від купруму можна пояснити тим, що формування правильного домену зв’язування індукується лише в тому випадку, коли декілька іонів купруму за допомогою хелатних зв’язків приєднуються до залишків цистеїну, розміщених в даній частині білкової молекули. Цікавим є те, що С-кінцева половина CITF має чітко виражений кислий характер, що притаманно й іншим факторам активації транскрипції [3].
Основні транскрипційні фактори
Завдяки біохімічному аналізу білків дріжджів та ссавців вдалось достатньо добре вивчити комплекс РНК-полімерази ІІ (Pol II) та механізми ініціації транскрипції.
У дріжджів необхідними компонентами для точної ініціації транскрипції in vitro є РНК-полімераза ІІ, що складається з 12 субодиниць, та основні транскрипційні фактори – TBP, TFIIE,TFIIB, TFIIF і TFIIH [6]. TBP зв’язується з ТАТА-елементом, TFIIВ займає ділянку між ТАТА-боксом і сайтом ініціації транскрипції, TFIIF взаємодіє з Pol II та відіграє роль у приєднані РНК-полімерази ІІ до промотора, TFIIH фосфорилює С-кінець найбільшої субодиниці Pol II і TFIIE здійснює очистку, усунення перешкод на промоторі. Крім перелічених, припускають також наявність TFIIА-фактора, що впливає на зв’язування TBP з ТАТА-елементом і є необхідними для точної ініціації in vitro. Виділяють також TBP-асоційовані фактори (TAFs (TBP associating factors) і компоненти голоферменту Pol II – Srb та інші білки, що, будучи основними транскрипційними факторами, однак, не потрібні для точної ініціації in vitro. Утворення активного транскрипційного комплексу може відбуватись in vitro, шляхом поступового додавання окремих факторів [5, 6]. Проте, існування великого комплексу, що вже містить основні транскрипційні фактори вказує на те, що весь транскрипційний апарат, за винятком TBP та TFIIB повністю сформований і функціональний перед зв’язуванням з промотором, а не утворюється на ньому.
ТАТА-зв’язуючий білок (TBP) – це найбільш консервативний еукаріотичний транскрипційний фактор, С-кінцевий core-домен якого складається з 180 амінокислот, більш ніж 80% послідовності яких гомологічна з іншими еукаріотичними видами. С-кінцевий core-домен TBP є необхідним і достатнім для нормального клітинного росту, тоді як еволюційно мінливий N-кінцевий домен функціонально несуттєвий. Проте, core-домен неповного TАТА-зв’язуючого білка значно інгібує клітинний ріст [5, 6].
Сore-домен – високомолекулярний димер споріднених, але не ідентичних субдоменів. До його складу входять α-спіралі та β-складчасті структури. При зв’язуванні з ТАТА-боксом виникає спотворення подвійної спіралі ДНК безпосередньо перед послідовністю впізнавання. Орієнтація TBP на ТАТА-елементі визначає локалізацію взаємодіючих компонентів активного механізму транскрипції і її напрямленість.
Було встановлено, що TBP виступає не лише транскрипційним фактором Pol II, а також є необхідним для всіх трьох ядерних РНК-полімераз. Така багатофункціональність TBP забезпечується асоціацією з іншими білками в окремі комплекси (SL1, TFIID, TFIIB), які є специфічними для РНК-полімераз. Як з’ясувалось, за відсутності TBP не проходить транскрипція жодного гену in vivo, навіть якщо ген втратив ТАТА-елемент.
Дріжджовий TBP може асоціюватись з Pol II-специфічними TAFs та утворювати комплекс, подібний до TFIID інших еукаріот. Дріжджовий TFIID здатний підтримувати базову та активовану транскрипцію in vitro, тоді як сам ТАТА-зв’язуючий білок не здатний взаємодіяти чи реагувати на білки-активатори. Всі відомі TAFs необхідні для підтримання нормальної життєздатності клітин. Проте, на даний час, не зрозуміло чим обумовлена така потреба у TBP-асоційованих факторах. Існують припущення, що TAFs відіграють важливу роль у транскрипції більшості генів, мають специфічний вплив на активацію чи репресію, або транскрипцію одного чи кількох дуже важливих генів-регуляторів [6].
Досі нез’ясовано чи транскрипція дріжджів з Pol II промоторів опосередкована через TBP чи TFIID, оскільки, на відміну від інших організмів дріжджові клітини містять значну кількість вільного TBP, а також було показано, що активована транскрипція може відновлюватись in vitro за відсутності TAFs і TFIID.
Дріжджовий TFIIА містить дві субодиниці, що є важливими для підтримання клітинної життєздатності. Зменшення активності TFIIА in vivo пригнічує активність кількох Pol II генів, проте не впливає на Pol IIІ транскрипцію. Ефективна взаємодія між TFIIА і TBP є важливою для транскрипційної активації in vivo.
TFIIВ і Pol II – транскрипційні фактори, що відповідають за вибір сайту ініціації. S cerevisiae відрізняється від інших еукаріотичних організмів більшою віддалю між ТАТА-елементом і сайтом ініціації та варіативністю цієї віддалі. In vivo, мутації TFIIВ гену чи гену найбільшої субодиниці Pol II (Rpb1) можуть пригнітити ініціацію з нормального сайту, та зумовити ініціацію транскрипції з сайтів, що знаходяться downstream. І навпаки, мутації іншої субодиниці Pol II – Rpb 9 – змінюють положення сайту upstream [6, 7].
TFIIН пов’язує Pol II транскрипцію, ексцизійну репарацію та контроль клітинного циклу. Цей фактор фосфорилює С-кінцевий хвіст найбільшої субодиниці Pol II. Складається з п’яти субодиниць. Значна кількість дріжджової РНК-полімерази ІІ знаходиться у великому білковому комплексі – Pol II голоензимі. In vitro Pol II голоензим реагує на активатори транскрипції. Деякі компоненти голоензиму є необхідними для клітинного росту, оскільки їх втрата веде до загальної втрати Pol II транскрипції.

