




Спробуємо тепер, виходячи із загальноекологічних міркувань про трансформацію речовин у системі і ґрунтуючись на асимптотичних оцінках, максимально спростити модель, щоб провести її аналітичне дослідження.
У блоці ”Консументи” поєднуються біомаси зоопланктону, бентосу, риб і т. ін., але навіть при такій агрегації в реальних екосистемах біомаса консументів є малою величиною порівняно з біомасами продуцентів або бактеріопланктону. Водночас у цьому блоці постійно відбувається переробка значних кількостей речовини: виїдається фітопланктон, виділяється детрит. З іншого боку, звичайно біомаса консументів – порівняно повільно змінний компонент екосистеми. Це пояснюється тим, що, по-перше, консументи пов’язані в ієрархічні трофічні ланцюги зі стійкою структурою, а по-друге – мають доволі складну трофічну поведінку, що дає їм змогу значно меншою мірою залежати від коливань ресурсу.
Усе це дає підстави розглядати консументи як проточний блок, в якому відбувається швидка трансформація речовини. Тому можна вважати, що qaz» qzs та dz / dt має порядок e, де e << 1 – малий параметр.
Тоді z (t)» z 0 = const, а
qaz = Va (a, z) z» Va (a, z 0) z 0,
де Va (a, z) – трофічна функція споживання фітопланктону консументами.
У природних системах трофічні ланцюги, як правило, є напруженими і ефект насичення відсутній. Якщо до того ж припустити, що внутрішньовидовою конкуренцією серед консументов можна знехтувати, то, Va (a, z) z природно записати у вигляді: Va (a, z 0) z 0» m 1 az 0 =
=  a. При цьому константа a описує певну додаткову «смертність» фітопланктону за рахунок консументів. Збільшення складності трофічних ланцюгів в екосистемі (як і збільшення біомаси консументів) призводить до зростання a.
a. При цьому константа a описує певну додаткову «смертність» фітопланктону за рахунок консументів. Збільшення складності трофічних ланцюгів в екосистемі (як і збільшення біомаси консументів) призводить до зростання a.
Оскільки характерні часи росту мікроорганізмів є меншими за характерні часи решти процесів, то можна вважати, що біомаса бактерій не лімітує швидкості біорозкладу органічної речовини: чисельність бактерій-деструкторів швидко підлаштовується до змін середовища, практично не затримуючи процес розкладання. Звичайно, це справджується тільки для доволі усереднених систем, в яких не враховуються різні ефекти температурного, світлового, токсичного і т. ін. інгибіювання.
З урахуванням ієрархії часу відповідні рівняння для бактерій в (1) точніше було б записати у вигляді:
e  = qsb – qbn.
= qsb – qbn.
Величина b (t) швидко прямує до свого стаціонарного значення, яке природно вважати функцією біомаси детриту: чим більшою є кількість мертвої органіки, тим більшою є біомаса бактерій, які її розкладають. Отже, при t ® ¥: b (t) ® b* (s)» l s.
Після цього можна вважати, що потік qsb приблизно дорівнює потоку qbn,оскільки біомаса детриту s у бактеріальному масштабі часу є повільною змінною, і тому db */ dt» 0. Потік qsb можна виразити в такому вигляді:
qsb = qsb (s, b, x) = m0(x) Q (s, b),
де m0(x) – кисневий коефіцієнт, Q (s, b) – швидкість мінералізації органіки.
При таких припущеннях Q (s, b)» Q (s, b *)» l0 s. Таким чином, qsb = l0m0(x) s = s m(x). Тут m(x) – коефіцієнт бактеріального розпаду детриту, який залежить від концентрації кисню у воді (рис. 2).

Рис. 2. Залежність швидкості бактеріального розкладання детриту від концентрації кисню у воді
Функція m(x) відображає зміну швидкості біорозкладу при перемиканні від аеробних до анаеробних умов:  і
і  – певні порогові значення концентрації О2; х > відповідає аеробним умовам, х < – анаеробним, а при < х < встановлюються певні проміжні режими.
– певні порогові значення концентрації О2; х > відповідає аеробним умовам, х < – анаеробним, а при < х < встановлюються певні проміжні режими.
Нехай qna = aVn (n, а), де Vn (n, а) – трофічна функція споживання біогенів фітопланктоном.
У найпростішій моделі вважається, що Vn = Vn (n). Оскільки Vn (0) = 0 і Vn (¥) < ¥, то для конкретного завдання Vn (n) можна використовувати функцію Моно:
Vn (n) =  ,
,
де a – максимальна швидкість росту фітопланктону, d – константа Міхаеліса.
Природна смертність фітопланктону задається у вигляді: qas = maa; тоді сумарна втрата біомаси фітопланктону:
qas + qaz = maa + a = r a.
Можна вважати, що потік кисню, утворюваного при фотосинтезі, лінійно залежить від біомаси фітопланктону: qaх = qaх (а) = b a, де b – коефіцієнт швидкості виділення кисню.
Нарешті, qхb = g qsb, де g – коефіцієнт споживання кисню при розкладі органічної речовини.
Збираючи всі зроблені правдоподібні припущення, можна звести модель (1) з до вигляду:
 =
= 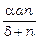 – r a;
– r a;  = – + s m(x);
= – + s m(x);
 = r a – s m(x); (1)
= r a – s m(x); (1)
 = k (xn – x) + b a – g s m(x) .
= k (xn – x) + b a – g s m(x) .

