





Аутосомное наследование. Характерные черты аутосомного наследования признаков обусловлены тем, что соответствующие гены, расположенные в аутосомах, представлены у всех особей вида в двойном наборе. Это означает, что любой организм получает такие гены от обоих родителей. В соответствии с законом чистоты гамет в ходе гаметогенеза все половые клетки получают по одному гену из каждой аллельной пары (рис. 6.6). Обоснованием этого закона является расхождение гомологичных хромосом, в которых располагаются аллельные гены, к разным полюсам клетки в анафазе I мейоза (см. рис. 5.6).
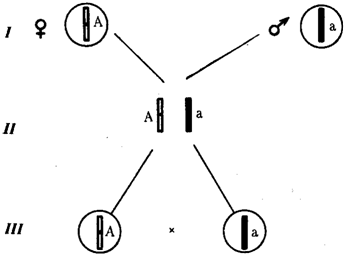
Рис. 6.6. Обоснование закономерностей аутосомного наследования признаков:
I — гаплоидные гаметы родителей, II — диплоидный генотип особи (фенотип зависит от взаимодействия аллельных генов Аа); III— гаплоидные гаметы гетерозиготной особи (гаметы «чисты», так как несут по одному из пары аллельных генов); черным и белым обозначены гомологичные хромосомы; буквами — определенные локусы
Ввиду того что развитие признака у особи зависит в первую очередь от взаимодействия аллельных генов, разные его варианты, определяемые разными аллелями соответствующего гена, могут наследоваться по аутосомно-доминантному или аутосомно-рецессивному типу, если имеет место доминирование. Возможен также промежуточный тип наследования признаков при других видах взаимодействия аллелей (см. разд. 3.6.5.2).
При доминировании признака, описанном Г. Менделем в его опытах на горохе, потомки от скрещивания двух гомозиготных родителей, различающихся по доминантному и рецессивному вариантам данного признака, одинаковы и похожи на одного из них (закон единообразия F1). Описанное Менделем расщепление по фенотипу в F2 в отношении 3:1 в действительности имеет место лишь при полном доминировании одного аллеля над другим, когда гетерозиготы фенотипически сходны с доминантными гомозиготами (закон расщепления в F2).

Рис. 6.7. Аутосомное наследование признака:
I — полное доминирование (наследование цвета лепестков у гороха); II — неполное доминирование (наследование цвета лепестков у ночной красавицы)
Наследование рецессивного варианта признака характеризуется тем, что он не проявляется у гибридов F1, а в F2 проявляется у четверти потомков (рис. 6.7.7).
В случаях формирования у гетерозигот нового варианта признака по сравнению с гомозиготами, что наблюдается при таких видах взаимодействия аллельных генов, как неполное доминирование, кодоминирование, межаллельная комплементация, гибриды F1 не похожи на родителей, а в F2 образуется три фенотипических группы потомков (рис. 6.7, II).
Завершая описание характерных черт аутосомно-доминантного и аутосомно-рецессивного наследования, уместно напомнить, что хотя в случае доминирования одного из аллелей присутствие в генотипе другого, рецессивного, аллеля не сказывается на формировании доминантного варианта признака, на фенотипическое проявление аллеля оказывает влияние вся система генотипа конкретного организма, а также среда, в которой реализуется наследственная информация. В связи с этим существует возможность неполной пенетрантности доминантного аллеля у особей, имеющих его в генотипе.

Рис. 6.8. Наследование признака окраски глаз у дрозофилы:
I, II — различие результатов скрещивания в зависимости от пола родителя с доминантным признаком; зачернены отцовские хромосомы
Сцепленное с полом наследование. Анализ наследования признака окраски глаз у дрозофилы в лаборатории Т. Моргана выявил некоторые особенности, заставившие выделить в качестве отдельного типа наследования признаков сцепленное с полом наследование (рис. 6.8).
Зависимость результатов эксперимента от того, кто из родителей являлся носителем доминантного варианта признака, позволила высказать предположение, что ген, определяющий окраску глаз у дрозофилы, расположен в Х-хромосоме и не имеет гомолога в Y-хромосоме. Все особенности сцепленного с полом наследования объясняются неодинаковой дозой соответствующих генов у представителей разного — гомо- и гетерогаметного пола.
Гомогаметный пол несет двойную дозу генов, расположенных в Х-хромосоме. Развитие соответствующих признаков у гетерозигот (ХAХa) зависит от характера взаимодействия между аллельными генами. Гетерогаметный пол имеет одну Х-хромосому (ХО или XY). У некоторых видов Y-хромосома генетически инертна, у других она содержит некоторое количество структурных генов, часть из которых гомологична генам Х-хромосомы (рис. 6.9). Гены негомологичных участков Х- и Y-хромосом (или единственной Х-хромосомы) у гетерогаметного пола находятся в гемизиготном состоянии. Они представлены единственной дозой: ХAY, ХaХ, XYB. Формирование таких признаков у гетерогаметного пола определяется тем, какой аллель данного гена присутствует в генотипе организма.
Характер наследования сцепленных с полом признаков в ряду поколений зависит от того, в какой хромосоме находится соответствующий ген. В связи с этим различают Х-сцепленное и Y-сцепленное (голандрическое) наследование.
Х-сцепленное наследование. Х-хромосома присутствует в кариотипе каждой особи, поэтому признаки, определяемые генами этой хромосомы, формируются у представителей как женского, так и мужского пола. Особи гомогаметного пола получают эти гены от обоих родителей и через свои гаметы передают их всем потомкам. Представители гетерогаметного пола получают единственную Х-хромосому от гомогаметного родителя и передают ее своему гомогаметному потомству.
У млекопитающих (в том числе и человека) мужской пол получает Х-сцепленные гены от матери и передает их дочерям. При этом мужской пол никогда не наследует отцовского Х-сцепленного признака и не передает его своим сыновьям (рис. 6.10).
Так как у гомогаметного пола признак развивается в результате взаимодействия аллельных генов, различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. Х-сцепленный доминантный признак (красный цвет глаз у дрозофилы) передается самкой всему потомству. Самец передает свой Х-сцепленный доминантный признак лишь самкам следующего поколения. Самки могут наследовать такой признак от обоих родителей, а самцы — только от матери.

Рис. 6.9. Схема гомологичных и негомологичных локусов
в половых хромосомах человека:
I — Х-хромосомы: заштрихованы локусы, отсутствующие в Y-хромосоме (красно-зеленая слепота, гемофилия и др.); II — Y-хромосома: заштрихованы локусы, отсутствующие в Х-хромосоме (перепонки между пальцами, гены-детерминаторы развития организма по мужскому типу); участки Х- и Y-хромосом, соответствующие гомологичным локусам, не заштрихованы

Рис. 6.10. Обоснование закономерностей сцепленного с полом
наследования признаков:
I — сочетание половых хромосом в кариотипе представителей разного пола; II — гомогаметный пол образует один тип гамет, гетерогаметный — два; III — представители гомогаметного пола получают хромосомы от обоих родителей; представители гетерогаметного пола получают Х-хромосому от гомогаметного родителя, а Y-хромосому — от гетерогаметного родителя; это справедливо для генов, расположенных в негомологичных локусах Х- и Y-хромосом; зачернены отцовские хромосомы
Х-сцепленный рецессивный признак, (белый цвет глаз у дрозофилы) у самок проявляется только при получении ими соответствующего аллеля от обоих родителей (XaXa). У самцов XaY он развивается при получении рецессивного аллеля от матери. Рецессивные самки передают рецессивный аллель потомкам любого пола, а рецессивные самцы —только «дочерям» (см. рис. 6.8).При Х-сцепленном наследовании, так же как и при аутосомном, возможен промежуточный характер проявления признака у гетерозигот. Например, у кошек пигментация шерсти контролируется Х-сцепленным геном, разные аллели которого определяют черную (XA и рыжую (XA’) пигментацию. Гетерозиготные самки XAXA’ имеют пеструю окраску шерсти. Самцы же могут быть либо черными (XAY, либо рыжими (XA’Y).

