




Биоэлектрические явления - "животное электричество" было открыто в 1791 г. итальянским ученым Гальвани. Данные современной мембранной теории происхождения биоэлектрических явлений получены Ходжкиным, Кацом и Хаксли в исследованиях, проведенных с гигантским нервным волокном кальмара (диаметром 1 мм) в 1952 г.
Плазматическая мембрана клетки (плазмолемма), ограничивающая снаружи цитоплазму клетки, имеет
толщину около 10 нм и состоит из двойного слоя липидов, в который погружены глобулы белков (молекулы, свернутые в клубки или спирали). Белки выполняют функции ферментов, рецепторов, транспортных систем, ионных каналов. Они либо частично, либо целиком погружены в липидный слой мембраны (рис. 13). В состав мембраны входит также небольшое количество углеводов.

Рис. 13. Модель клеточной мембраны как жидкой мозаики из липидов и белков - поперечный разрез (Стерки П., 1984). а - липиды; в - белки
Сквозь мембрану движутся различные вещества в клетку и из клетки. Регуляция этого процесса - одна из основных функций мембраны. Основными ее свойствами являются избирательная и изменчивая проницаемость. Для одних веществ она служит барьером, для других - входными воротами. Вещества могут проходить через мембрану по закону концентрационного градиента (диффузия от большей концентрации к меньшей), по электрохимическому градиенту (разная концентрация заряженных ионов), путем активного транспорта - работа натрий-калиевых насосов.
Мембранный потенциал, или потенциал покоя. Между наружной поверхностью клетки и ее цитоплазмой существует разность потенциалов порядка 60 - 90 мВ (милливольт), называемая мембранным потенциалом, или потенциалом покоя. Его можно обнаружить при помощи микроэлектродной методики. Микроэлектрод представляет собой тончайший стеклянный капилляр с диаметром кончика 0,2 - 0,5 мкм. Его заполняют раствором электролита (КС1). Второй электрод обычных размеров погружают в раствор Рингера, в котором находится исследуемый объект. Через усилитель биопотенциалов электроды подводят к осциллографу. Если под микроскопом с помощью микроманипулятора микроэлектрод ввести внутрь нервной клетки, нервного или мышечного волокна, то в момент прокола осциллограф покажет разность потенциалов - потенциал покоя (рис. 14). Микроэлектрод настолько тонок, что он практически не повреждает мембраны.
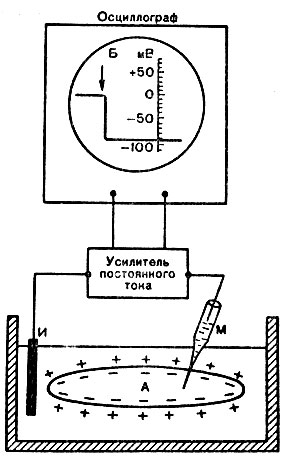
Рис. 14. Измерение потенциала покоя мышечного волокна (А) при помощи внутриклеточного микроэлектрода (схема). М - микроэлектрод; И - индифферентный электрод. Луч на экране осциллографа показан стрелкой
Мембрайно-ионная теория объясняет происхождение потенциала покоя неодинаковой концентрацией несущих электрические заряды К+, Na+ и Сl- внутри и вне клетки и различной проницаемостью для них мембраны.
В клетке в 30 - 50 раз больше К+ и в 8 - 10 раз меньше Na+, чем в тканевой жидкости. Следовательно, внутри клетки преобладают К+, снаружи - Na+. Основным анионом тканевой жидкости является Сl-. В клетке преобладают крупные органические анионы, которые не могут диффундировать сквозь мембрану. (Как известно, катионы имеют положительный заряд, а анионы - отрицательный.) Состояние неодинаковой ионной концентрации по обе стороны плазматической мембраны называют ионной асимметрией. Она поддерживается работой натрий-калиевых насосов, которые непрерывно перекачивают Na+ из клетки и К+ в клетку. Работа эта осуществляется с затратой энергии, освобождающейся при расщеплении аденозинтрифосфорной кислоты. Ионная асимметрия - физиологическое явление, сохраняющееся пока клетка жива.
В покое проницаемость мембраны значительно выше для К+, чем для Na+. В силу высокой концентрации ионы К+ стремятся выйти из клетки наружу. Сквозь мембрану они проникают на наружную поверхность клетки, но дальше уйти не могут. Крупные анионы клетки, для которых мембрана непроницаема, не могут последовать за калием, и скапливаются на внутренней поверхности мембраны, создавая здесь отрицательный заряд, который удерживает электростатической связью проскочившие через мембрану положительно заряженные ионы калия. Таким образом возникает поляризация мембраны, потенциал покоя; по обе ее стороны образуется двойной электрический слой: снаружи из положительно заряженных ионов К+, а внутри из отрицательно заряженных различных крупных анионов.
Потенциал действия. Потенциал покоя сохраняется до тех пор, пока не возникло возбуждение. Под действием раздражителя проницаемость мембраны для Na+ повышается. Концентрация Na+ снаружи клетки в 10 раз больше, чем внутри нее. Поэтому Na+сначала медленно, а затем лавинообразно устремляются внутрь. Ионы натрия заряжены положительно, поэтому происходит перезарядка мембраны и ее внутренняя поверхность приобретает положительный заряд, а наружная - отрицательный. Таким образом происходит реверсия потенциала, изменение его на обратный знак. Он становится отрицательным снаружи и положительным внутри клетки. Этим объясняется давно известный факт, что возбужденный участок становится электроотрицательным по отношению к находящемуся в покое. Однако повышение проницаемости мембраны для Na+ длится недолго; она быстро снижается и повышается для К+. Это вызывает усиление потока положительно заряженных ионов из клетки во внешний раствор. В итоге происходит реполяризация мембраны, ее наружная поверхность приобретает снова положительный заряд, а внутренняя - отрицательный.
Электрические изменения мембраны в процессе возбуждения получили название потенциала действия. Длительность его измеряется тысячными долями секунды (миллисекундами), амплитуда равна 90 - 120 мВ.
Во время возбуждения Na+ входят в клетку, а К+ выходят наружу. Казалось бы, что концентрация ионов в клетке должна меняться. Как показали опыты, даже многочасовое раздражение нерва и возникновение в нем десятков тысяч импульсов не изменяют содержания в нем Na+ и К+. Это объясняется работой натрий-калиевого насоса, который после каждого цикла возбуждения разводит ионы по местам: накачивает К+ обратно в клетку и выводит из нее Na+. Насос работает на энергии внутриклеточного обмена веществ. Это доказывается тем, что яды, прекращающие обмен веществ, прекращают работу насоса.
Потенциал действия, возникая в возбужденном участке, становится раздражителем для соседнего невозбужденного участка мышечного или нервного волокна и обеспечивает проведение возбуждения вдоль мышцы или нерва.
Возбудимость различных тканей неодинакова. Наиболее высокой возбудимостью отличаются рецепторы, специализированные структуры, приспособленные к улавливанию изменений во внешней среде и внутренней среде организма. Затем следует нервная, мышечная и железистая ткани.
Мерой возбудимости является порог раздражения, т. е. та наименьшая сила раздражителя, которая способна вызвать возбуждение. Порог раздражения иначе называют реобазой. Чем выше возбудимость ткани, тем меньшей силы раздражитель способен вызвать возбуждение.
Кроме того, возбудимость можно характеризовать тем временем, в течение которого должен действовать раздражитель, чтобы вызвать возбуждение, иначе говоря, порогом времени. Наименьшее время, в течение которого должен действовать электрический ток пороговой силы, чтобы вызвать возбуждение, называется полезным временем. Полезное время характеризует скорость течения процесса возбуждения.
Возбудимость тканей увеличивается в процессе умеренной деятельности и снижается при утомлении. Возбудимость претерпевает фазовые изменения во время возбуждения. Как только в возбудимой ткани возникает процесс возбуждения, она утрачивает способность отвечать на новое, даже сильное раздражение. Это состояние называется абсолютной невозбудимостью, или абсолютной рефрактерной фазой. Через некоторое время возбудимость начинает восстанавливаться. На пороговое раздражение ткань еще не отвечает, но на сильное раздражение отвечает возбуждением, хотя амплитуда возникающего потенциала действия в это время значительно снижена, т. е. процесс возбуждения слаб. Это фаза относительной рефрактерности. После нее возникает фаза повышенной возбудимости или супернормальности. В это время можно вызвать возбуждение очень слабым раздражителем, ниже пороговой силы. Только после этого возбудимость приходит в норму.
Для исследования состояния возбудимости мышечной или нервной ткани наносят два раздражения друг за другом через определенные интервалы. Первое вызывает возбуждение, а второе - тестирующее - испытывает возбудимость. Если на второе раздражение реакции нет, значит, ткань невозбудима; реакция слабая - возбудимость понижена; реакция усилена - возбудимость повышена. Так, если на сердце наносить раздражение во время систолы, то возбуждения не последует, к концу диастолы раздражение вызывает внеочередное сокращение - экстрасистолу, что свидетельствует о восстановлении возбудимости.
На рис. 15 сопоставлены во времени процесс возбуждения, выражением которого служит потенциал действия, и фазовые изменения возбудимости. Видно, что абсолютная рефрактерная фаза соответствует восходящей части пика - деполяризации, фаза относительной рефрактерности - нисходящей части пика - реполяризации мембраны и фаза повышенной возбудимости - отрицательному следовому потенциалу.

Рис. 15. Схемы изменений потенциала действия (а) и возбудимости нервного волокна (б) в различные фазы потенциала действия. 1 - местный процесс; 2 - фаза деполяризации; 3 - фаза реполяризации. Пунктиром на рисунке обозначены потенциал покоя и исходный уровень возбудимости

