Поиск:
Рекомендуем:
 Почему я выбрал профессую экономиста
Почему я выбрал профессую экономиста
 Почему одни успешнее, чем другие
Почему одни успешнее, чем другие
 Периферийные устройства ЭВМ
Периферийные устройства ЭВМ
 Нейроглия (или проще глия, глиальные клетки)
Нейроглия (или проще глия, глиальные клетки)
Категории:
Глава 5. Развитие глазного яблока 2 страница
|
|
|
|
|
Поиск: Рекомендуем:
Почему я выбрал профессую экономиста
Почему одни успешнее, чем другие
Периферийные устройства ЭВМ
Нейроглия (или проще глия, глиальные клетки)
Категории:
|
Глава 5. Развитие глазного яблока 2 страница
| Клетки нейрального гребня | Поверхностная эктодерма | ||
| Эктодерма | |||||
| Сенсорная часть сетчатки | Строма роговой оболочки и эндотелий. | Эпителий, железы и рес- | |||
| Склера (см. также мезодерма). | ницы кожи век и слез- | ||||
| Трабекулярная сеть | ного мясца | ||||
| Пигментный эпителий сет- | Оболочка и сухожилия наружных мышц глаза. | Эпителий конъюнктивы. | |||
| чатки | Соединительная ткань радужной оболочки | Хрусталик | |||
| Пигментный эпителий рес- | Мышцы ресничного тела. | Слезная железа. | |||
| ничного тела | Строма сосудистой оболочки | Слезоотводящая система | |||
| Непигментированный эпи- | Меланоциты (увеального тракта и кожи). | Стекловидное тело | |||
| телий ресничного тела | Оболочки зрительного нерва. | ||||
| Шванновские клетки ресничных нервов | |||||
| Пигментный эпителий ра- | Цилиарный ганглий. | ||||
| дужной оболочки | Кости глазницы. | ||||
| Хрящ | |||||
| Сфинктер и дилятатор ра- | Соединительная ткань глазницы. | ||||
| дужной оболочки | Мышечный слой и соединительнотканные оболочки | ||||
| кровеносных сосудов глазницы | |||||
| Зрительный нерв, аксоны | |||||
| и глия | |||||
| Стекловидное тело | |||||
| Мезодерма | |||||
| Волокна наружных мышц глаза. | |||||
| Эндотелиальная выстилка всех сосудов глазницы | |||||
| и глаза. | |||||
| Темпоральная часть склеры. | |||||
| Стекловидное тело |
Развитие роговой оболочки и склеры
573

|
|
|
|
|
|
|
Рис. 5.2.1. Схема развития роговой оболочки (центральные участки):
а — 39 дней развития. Двухслойный эпителий располагается на базальной мембране и отделен от трехслойного эндотелия узким межклеточным пространством; б — 7 недель развития. Мезенхима — будущая строма роговицы — перемещается в пространство между передним и задним эпителием; в — 7,5 недели развития. Мезенхима (фибробласты) формируют 5 слоев. Появляются коллагеновые волокна; г — к 3-му месяцу развития передний эпителий состоит из 2—3 слоев клеток и приблизительно 25—30 слоев стромальных фибробластов (кератоблас-
ружную стенку зрительного пузырька [171]. Образуется зрительный бокал. На определенном этапе погружения хрусталикового пузырька слой поверхностной эктодермы над ним смыкается и образуется зачаток роговицы. При этом роговица представляет собой довольно толстый слой эпителиальных клеток наружной эктодермы, расположенных на базальной мембране (рис. 5.1.11, 5.2.1—5.2.4). Базальные мембраны хрусталика и эпителия роговой оболочки на этом этапе развития временно «сплавлены». Несколько позже они разъединятся.
Между нейроэктодермальными краями зрительной чаши и хрусталиковым пузырьком начинается накопление недифференцированных клеток нейрального гребня (эктомезенхима), располагающихся спереди и по периферии хрусталика, а также по краям зрительной чаши.

Рис. 5.2.2. Последовательность миграции клеток экто-мезенхимы в передний отдел глазного яблока:
/—первая волна миграции, приводящая к формированию эндотелия роговой оболочки; //—вторая волна миграции с образованием радужной оболочки и части зрачковой мембраны; /// — третья волна миграции, завершающаяся образованием кератоци-тов стромы роговой оболочки (/—передний эпителий роговой оболочки; 2 — хрусталик; 3 — сетчатая оболочка)
тов). Появляются признаки начала формирования десцеметовой оболочки; д — 4,5 месяца развития. Отмечается дифференциация клеток переднего эпителия по слоям (появление крыловидных клеток). Четко видна базальная мембрана и боуменова оболочка. Строма в передних слоях еще нечетко организована; е — 7 месяцев развития. Строение роговой оболочки практически полностью соответствует строению роговицы взрослого. Отличием является лишь некоторая нечеткая организация передних слоев стромы
Определяется три волны миграции этих клеток (рис. 5.2.2). Первая волна приводит к накоплению клеток, которые в последующем дифференцируются в эндотелий роговой оболочки. Вторая волна обеспечивает закладку будущей стромы. Третья волна поставляет материал для формирования зрачковой мембраны и стромы радужной оболочки.
Изучение эмбриона цыпленка показало, что отделение хрусталикового пузырька стимулирует эпителиальные клетки базального слоя секретировать коллагеновые волокона и гли-козаминогликаны, выполняющие пространство между хрусталиком и эпителием роговицы. Это пространство составляет первичную стро- му роговицы. Первичная строма состоит из нежных микрофиламентов, аморфного материала и небольшого числа коллагеновых волокон. Из клеточных элементов она содержит только макрофаги.
Мезенхимные клетки, исходящие из нейрального гребня, расположены по краям зрительного бокала. Затем они мигрируют глубже базальной пластинки и формируют эндотелий роговицы. Эти клетки используют заднюю поверхность первичной стромы будущей роговой оболочки и базальную пластинку клеток хрусталика как субстрат для своего перемещения. Это передвижение клеток нервного гребня представляет первую волну из трех последовательных волн врастания клеток нейрального гребня в переднюю часть глаза.
Развитие эпителия. Приблизительно к 40-му дню эмбрионального развития (эмбрион 17 —18 мм) роговица состоит из поверхностного слоя плоских клеток, базального слоя кубовидных клеток, первичной стромы и двух слоев
Глава 5. РАЗВИТИЕ ГЛАЗНОГО ЯБЛОКА

|
|
|
| р •«_ -w~j. в»-"** Ч^ "^ |
«Г 8 г
Рис. 5.2.3. Эмбриональное развитие роговой оболочки (световая микроскопия):
а — строение роговой оболочки на 45 сутки (18 мм); б — строение роговой оболочки спустя 2 месяца (3! мм) эмбрионального развития: в — строение роговой оболочки спустя 5 месяцев эмбрионального развития (148 мм); г — строение передних слоев роговой оболочки новорожденного (51 см) (/ — передний эпителий: 2 — боуменова оболочка; 3 — строма; -/—задний эпителий
(эндотелий); 5 — хрусталик)
плоских эндотелиальных клеток, расположенных на ее задней поверхности (рис. 5.2.1—5.2.3).
К седьмой неделе беременности эпителиальные клетки поверхностного слоя имеют гексагональную форму. Их максимальный диаметр равняется 10 мкм [217—222, 291]. На апикальной поверхности клеток уже видны микроворсинки и складки. Клетки обладают довольно электронноплотной цитоплазмой, содержащей многочисленные гранулы гликогена. Поверхностно расположенные клетки соединяются между собой и клетками, лежащими глубже, при помощи единичных десмосом.
Между вторым и третьим месяцами эмбрионального развития большинство поверхностных клеток увеличивается в диаметре и достигает 15—20 мкм. Вся их апикальная поверхность уже покрыта многочисленными микроворсинками. Цитоплазма клеток несколько просвет-левает, а количество зерен гликогена уменьшается. Увеличивается и количество десмосом.
В начале четвертого месяца различается три типа поверхностных клеток. Это самые маленькие клетки, отличающиеся наличием большого количества микроворсинок и микроскладок, клетки среднего размера с промежуточным количеством микроворсинок и крупные клетки. Крупные клетки отличаются наименьшей элек-тронноплотностью цитоплазмы и находятся в состоянии десквамации, т. е. слущивания.
К 5—6-му месяцу эмбрионального развития передний эпителий роговицы приближается по строению к эпителию взрослых людей.
Развитие стромы. Развитию стромы роговой оболочки предшествует вторая волна миграции мезенхимных клеток нервного гребня в область «губы» зрительного бокала (рис. 5.2.2, 5.2.4). Эта миграция происходит в двух направлени-

Рис. 5.2.4. Ультраструктурные особенности боумено-вой оболочки на 7-м месяце эмбрионального развития. Четко определяется базальная мембрана эпителиоци-тов переднего эпителия (тонкие стрелки) и полу-десмосомы (широкая стрелка):
I — эпителиальная клетка; 2 — гранулы гликогена; 3 — боуменова оболочка; 4 — строма роговой оболочки
ях. На стадии развития эмбриона, достигшего 19 мм, клетки растут в пространство между эпителием хрусталика и эндотелием роговой оболочки и формируют первичную зрачковую мембрану. Примерно в это время основное вещество стромы роговой оболочки оттекает в результате гидратации гиалуроновой кислоты. Это способствует наступлению следующей волны миграции клеток нейрального гребня. На 7-й неделе эмбрионального развития (эмбрион 22—24 мм) мезенхимные клетки мигрируют и распределяются между эпителием роговой оболочки и эндотелием.
Ртвитие роговой оболочки и склеры
575

|
| / к Ш, |
Центральная область стромы первоначально не содержит клеток. Врастающие клетки дифференцируются в направлении стромальных фибробластов или кератоцитов, активно секре-тирующих коллагеновые волокна 1-го типа и основное вещество стромы.
Первоначально строма богата фибронекти-ном. Количество фибронектина постепенно уменьшается по мере увеличения числа мезен-химных клеток. Строма вскоре достигает своей максимальной толщины, которая приблизительно вдвое больше толщины постэмбриональной роговицы. Это происходит в результате уменьшения ее гидратации, уменьшения содержания гиалуроновой кислоты, а также ретракции (сокращения) коллагеновых волокон. Звездчатые мезенхимные клетки, беспорядочно рассеянные в строме, постепенно приобретают веретено-видную форму и ориентируются параллельно поверхности роговой оболочки. Это морфоге-нетическая трансформация начинается в задних слоях роговицы и сопровождается быстрым появлением коллагеновых волокон. Перед появлением волокон отмечается накопление гликозаминогликанов, богатых карбоксильными и сульфатными остатками. Кератансульфат не обнаруживается до 6-го месяца беременности [12]. В последующем коллагеновые волокна образуют пластинки. Пластинки удлиняются и утолщаются в результате постепенного накопления коллагеновых волокон (внутритканевой рост).
К 8-й неделе развития (30 мм) строма роговой оболочки в центральных участках состоит из пяти—восьми рядов клеток. Периферические участки стромы сливаются с мезенхимной тканью, которая в последующем превращается в склеру. Количество слоев стромы быстро увеличивается и на стадии развития, соответствующей 35 мм длины эмбриона, состоит из 15 слоев клеток и небольшого количества коллагеновых волокон. Задняя поверхность стромы покрыта эндотелием, состоящим из двух слоев клеток.
Как было указано выше, коллагеновые волокна растут в длину, в связи с чем увеличивается диаметр роговицы. Диаметр роговой оболочки эмбриона человека на 12-й неделе развития равен 2 мм, на 15-й неделе — 3,5 мм, на 17-й неделе — 4,5 мм, на 21-й неделе — 5,5 мм и на 35-й неделе — 9,3 мм [62]. Как диаметр роговой оболочки, так и ее толщина определяется постоянным взаимодействием роговой оболочки с формирующейся сетчаткой и хрусталиком [42, 137].
Развитие десцеметовой мембраны и эндо телия (рис. 5.2.5). К третьему месяцу развития (эмбрион 63 мм) эндотелий в центральной области роговицы становится однослойным и плоским. Располагается он на прерывистой ба-зальной пластинке, будущей десцеметовой мембране. На этой стадии развития базальная пла-
Рис. 5.2.5. Развитие десцеметовой оболочки (электронная микроскопия). Стрелками указана динамика изменения толщины десцеметовой оболочки. При этом увеличивается объем волокнистого материала оболочки:
а — 3-я неделя эмбрионального развития; б— 12-я неделя развития; в — 6-й месяц эмбриогенеза; г — момент рождения (/ — строма роговой оболочки; 2 — десцеметова оболочка; 3 — эндотелий роговицы)
стинка состоит из электроннопрозрачной зоны (lamina lucida, толщина — 37,5 нм), смежной с эндотелиальными клетками, и электронноплот-ной зоны (lamina densa, толщина — 36,7 нм), прилежащей к строме роговицы [152, 154, 282, 284, 286]. Дальнейшая дифференциация и утолщение десцеметовой мембраны происходят благодаря секреции эндотелиальными клетками «мембраноподобных структур». Процесс протекает довольно быстро. Если на 12-й неделе развития обнаруживается всего один слой, то к шестому месяцу слоев уже десять, а на момент рождения «мембраноподобных структур» тридцать—сорок. Параллельно с накоплением стромальных пластин появляются микрофиламен-ты длиной 170 нм и диаметром 40 нм. Эти филаменты располагаются перпендикулярно слоям стромальных пластин и связывают стро-мальные пластины, прикрепляя их к мембране. Именно благодаря этим филаментам происходит уплотнение области будущей десцеметовой мембраны [154].
Фибриллы будущей десцеметовой мембраны формируют гексагональные структуры, волокна в которых связаны «узлами» электронноплот-ного материала. В результате этого образуется равносторонний треугольник, стороны которого равны приблизительно ПО нм.
576
Глава 5. РАЗВИТИЕ ГЛАЗНОГО ЯБЛОКА
 К четвертому месяцу эмбрионального развития апикальные поверхности эндотелиаль-ных клеток соединяются при помощи десмосом. Образование контактов совпадает с началом секреции ресничным телом камерной влаги.
К четвертому месяцу эмбрионального развития апикальные поверхности эндотелиаль-ных клеток соединяются при помощи десмосом. Образование контактов совпадает с началом секреции ресничным телом камерной влаги.
К шестому месяцу десцеметова мембрана уже четко различается при микроскопическом исследовании. При этом строма роговицы состоит из основного вещества, коллагеновых волокон и многочисленных активных кератоцитов [262].
Максимальной толщины десцеметова мембрана достигает на момент рождения.
Развитие боуменовой оболочки. На 18-й неделе эмбрионального развития базальная пластинка эпителия роговицы состоит из двух слоев — относительно тонкой lamina lucida (толщина— 41,68 нм) и полностью развитой la mina densa. Причем между эпителиальными клетками и базальной пластинкой полудесмо-сомы и «якорные» микрофиламенты развиты лишь частично. Постепенно микрофиламенты (средняя длина — 474 нм и диаметр — 17,6 нм) возникают у базальной поверхности эпителиальных клеток, пересекают lamina densa и lu cida и заканчиваются в боуменовом слое [10].
Формируется боуменова оболочка в результате синтетической активности фибробластов передних слоев будущей стромы роговой оболочки. Структурные компоненты оболочки (основное вещество, волокна) также способны синтезировать клетки переднего эпителия роговицы [179].
К 26-й неделе беременности вблизи полудес-мосом уже обнаруживаются «якорные» микрофиламенты типичного строения (рис. 5.2.4).
Необходимо отметить, что роговая оболочка эмбриона полупрозрачна, поскольку она поглощает воду в значительно большем количестве, чем роговица взрослого. Степень гидратации роговицы постепенно уменьшается по мере вызревания переднего и заднего эпителия, и на момент рождения она прозрачна. Нервные волокна в строме роговицы можно обнаружить уже на 3-м месяце развития, а к 5-му месяцу нервные окончания уже довольно равномерно распределены в эпителии [125, 172, 275]. Электронномикроскопические исследования выявили, что созревание синаптических образований нервных окончаний роговой оболочки также происходит на пятом месяце развития.
Развитие склеры. Источником развития склеры являются клетки мезенхимы, которые постепенно уплотняются вокруг зрительного бокала (рис. 5.1.11). Исходят эти клетки, в основном, из нейрального гребешка [14, 262]. У млекопитающих задняя часть склеры, вероятно, развивается из парааксиальной мезодермы [162—165], особенно в местах прикрепления наружных мышц глаза.
Развитие склеры начинается в передних отделах на 6,5 недели развития эмбриона, и про-
цесс постепенно распространяется назад. На этой стадии определяется восемь—девять параллельно расположенных клеточных слоев (фибробласты) [218]. Позади экватора клетки располагаются более беспорядочно.
Цитоплазма клеток содержит многочисленные свободные рибосомы, небольшое количество шероховатого эндоплазматического ретику-лума с хлопьевидным материалом. Обнаруживается и плохо развитый аппарат Гольджи.
Гранулы гликогена и капли липидов, являющиеся источником энергии, более многочисленны в клетках, расположенных в переднем отделе будущей склеры. В межклеточном пространстве можно обнаружить беспорядочно распределенные коллагеновые фибриллы (диаметр — 27—29 нм). Эластические волокона на ранних стадиях развития склеры не обнаруживаются.
К середине седьмой недели эмбрионального развития уже четко определяется граница между склерой и окружающими тканями. Число клеточных слоев в передних отделах достигает 15. Клетки, прилежащие к сосудистой оболочке, отличаются большей длиной и более компактным расположением. Шероховатая эндо-плазматическая сеть склеральных клеток хорошо развита. Определяется аппарат Гольджи, вблизи которого видны многочисленные зерна гликогена и капельки липидов.
Количество коллагеновых волокон к этому моменту также увеличивается. При этом их диаметр увеличивается до 30—40 нм. Появляются первые отложения эластина в виде микрофибрилл диаметром 10—12 нм.
По мере накопления межклеточного вещества исчезают контакты между цитоплазматичес-кими отростками соседних склеральных клеток.
В течение девятой недели эмбрионального развития фибробласты задней части склеры отдают длинные отростки, ориентирующиеся параллельно слою хориокапилляров сосудистой оболочки. Нарастает количество эластических волокон.
К одиннадцатой неделе (60—65 мм) существенных различий в строении внутренних и наружных слоев склеры уже не выявляется. В это время часть мезенхимных клеток распространяется между нервными волокнами зрительного нерва, они ориентируются поперечно и формируют решетчатую пластинку.
В начале четвертого месяца склера в пре- и постэкваториальных областях состоит из 30 слоев клеток, а к шестому месяцу — из 50 слоев. В дальнейшем количество слоев не увеличивается [219]. Увеличивается лишь объем межклеточного вещества и количество коллагеновых (диаметр — 100 нм) и эластических волокон. Необходимо отметить, что, хотя клеточные элементы роговой оболочки и склеры имеют общее происхождение (мезенхима), они ведут себя по-разному в отношении синтетической активности. Herrmann [101] установил, что
Развитие роговой оболочки и склеры
577
синтезируемый роговичными клетками коллаген отличается по своему составу от коллагена, синтезируемого клетками склеры. Более того, различно соотношение коллагеновых и неколла-геновых белков и скорость их синтеза.
Несколько позже (эмбрион 70 мм) во внутреннем слое передней части склеры развивается клиновидной формы выступ, склеральная шпора. Форма этой структуры объясняется натяжением участка склеры развивающимися волокнами ресничной мышцы. К середине седьмого месяца передние концы волокон продольной ресничной мышцы прикрепляются к склеральной шпоре. Коллагеновые и эластические волокна в области склеральной шпоры ориентированы циркулярно [262].
Интрасклеральные сосудистые сплетения, водяные вены и коллекторные каналы, пересекающие склеру в лимбальной области, впервые появляются на 12-й неделе эмбрионального развития. Эти сосуды дифференцируются из примитивной мезодермы.
Поскольку склера определяет размер и форму глазного яблока, факторы, регулирующие ее развитие, играют основную роль в формировании рефракционных особенностей глаза. Факторов, индуцирующих развитие слеры, как и других структур глаза, большое количество, и сек-ретируются они многими структурами. Одним из таких факторов, принимающих особое участие в дифференциации склеры, является матричная РНК, контролирующая синтез инсулин-подобного фактора роста. В наибольшем количестве обнаруживается он в клетках нейраль-
ного гребня, откуда и направляется к развивающимся структурам [87]. В зрелой склере, роговице и сетчатке эта РНК не выявляется [42]. Дальнейший рост склеры после рождения контролируется другими факторами роста, которые синтезируются и высвобождаются из сетчатки. Сигнал, побуждающий к развитию склеры, относится к пептидным гормонам, синтез и выделение которых стимулируется светом, попадающим на развивающуюся сетчатку. Именно по этой причине у новорожденных, лишенных, по разным причинам, зрительных стимулов, отмечается неполное созревание склеры, что может явиться причиной развития близорукости.
5.3. РАЗВИТИЕ ПЕРЕДНЕЙ КАМЕРЫ И ДРЕНАЖНОЙ СИСТЕМЫ
Началом формирования передней камеры глаза является образование щелевидного пространства, возникающего в результате миграции в эту область первой волны (будущего эндотелия роговицы) и второй волны (первичная зрачковая мембрана) мезенхимных клеток [6, 14]. Дальнейшее развитие передней камеры сводится к дифференциации структур, ограничивающих это пространство (рис. 5.2.2).
Развитие угла передней камеры. Приблизительно на 7-й неделе эмбрионального развития (эмбрион 22—24 мм) угол передней камеры выполнен свободно лежащими мезенхимны-ми клетками, участвующими в формировании трабекулярной сети (рис. 5.3.1; рис. 5.3.2, а).
|
|
Рис. 5.3.1. Схема формирования угла передней камеры и отношение его к соседним структурам:
а —3-й месяц эмбрионального развития. Эндотелий роговой оболочки распространяется на угол передней камеры. Отмечается начало формирования шлеммова канала (стрелка), а также конденсация соединительной ткани с образованием склеральной шпоры (полая стрелка). По передней поверхности переднего листка глазного бокала (будущий пигметныи эпителий радужной оболочки) развиваются кровеносные сосуды; б — 4-й месяц эмбрионального развития. Определяется углубление будущего угла передней камеры глаза, эндотелий в этой области исчезает. Четко виден шлеммов канал (стрелка). От внутреннего листка губы глазного бокала в направлении капсулы хрусталика направляются волокна (будущий зонулярный аппарат); в — 5-й месяц эмбрионального развития. Сформирована радужная оболочка. Угол передней камеры выполнен пучками соединительной ткани, между которыми определяются свободные пространства. Начало форми-
рования дилятатора радужки. Стрелка указывает на сосуд большого круга кровообращения радужки; г — 5-й месяц развития. Зрачковый край радужки. Сформирован сфинктер. Поверхностные слои стромы радужки содержат сосуды; д — 6-й месяц эмбрионального развития. Схема развития дилятатора радужки. В это время апикальные поверхности нейроэпителиальных клеток вытягиваются, направляясь в строму радужки. В них появляются мио-филаменты (/—угол передней камеры; 2 — роговая оболочка; 3 — эндотелий роговой оболочки; 4 — зрачковая мембрана; 5 — наружный нейроэпителиальный слой губы глазного бокала; 6 — внутренний нейроэпителиальный слой губы глазного бокала; 7 — хрусталик; 8 — клетки дилятатора радужной оболочки; 9 — сосуды большого круга кровообращения радужки; 10 — сосуды малого круга кровообращения радужки; // — волокна будущего зонуляр-ного аппарата; 12 — структурные элементы дренажной системы)
578
Глава 5. РАЗВИТИЕ ГЛАЗНОГО ЯБЛОКА

|
|
|
|
|
|
|
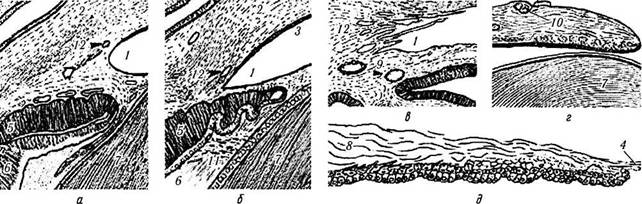
Рис. 5.3.2. Развитие угла передней камеры (световая микроскопия):
а — эмбрион 6.4 недели (по Sellheyer, Spitznas, 1988). Мезенхима, распложенная вокруг глазного бокала, конденсируется с образованием слоев клеток. Этот процесс характеризует начало формирования склеры (/ — мезенхимные клетки; 2 — эмбриональная конъюнктива; 3 —внутренний слой глазного бокала; 4 — наружный слой глазного бокала; 5 — передние участки стекловидного тела; 6 — хрусталик); б — эмбрион на 11-й неделе развития (по Ozanics, Jakobiec, 1982). Наружный слой глазного бокала пигментирован и волнистый. В мезенхиме на границе с наружным нейроэпителиальным слоем формируются сосуды. Мезенхима распространяется на переднюю поверхность хрусталика. В результате этого процесса формируется эмбриональная передняя камера (/—роговая оболочка; 2 —передняя камера; 3 —зрачковая мембрана; 4 — капсулопупиллярные сосуды гиалоидной системы; 5 — стекловидное тело); в — плод на четвертом месяце развития.
На задней поверхности будущего угла располагаются клетки, из которых в последующем развивается сосудистая сеть зрачковой мембраны. Кроме того, можно обнаружить свободно лежащие мезенхимные клетки и врастающие в зрительный бокал клетки пигментного эпителия. Эндотелиальные клетки распространяются на угол, а к 15-й неделе покрывают переднюю поверхность будущей радужки. С этого момента очерчивается угол передней камеры (рис. 5.3.1; рис. 5.3.2, б). До сих пор неизвестны непосредственные механизмы формирования угла передней камеры в этот период времени. Извест-
Пигментированный наружный и беспигментный внутренний слои края глазного бокала формируют отростки ресничного тела, которые лежат на уровне угла передней камеры. Между хрусталиком и отростками ресничного тела располагаются фибриллы эмбриональной цинновой связки (/ — угол передней камеры; 2 — часть края глазного бокала, относящаяся к будущей радужке; 3 — эмбриональная циннова связка; 4 — эпителий хрусталика; 5 — краевой синус; 6 — большой артериальный круг; 7— сетчатка; 8 — стекловидное тело (стрелкой указано место образования основания стекловидного тела; закругленной стрелкой указано место образования ресничной мышцы)); г — плод на 6-м месяце развития. Четко определяется трабекулярная сеть, шлеммов канал, склеральная шпора (/ — шлеммов канал; 2 — склеральная шпора; 3 — трабекулярная сеть; 4 — ресничная мышца; 5 — роговая оболочка; б — ресничные отростки; 7 — строма радужки)
но только, что углубление угла связано с различной скоростью роста, обратного развития и дифференциации мезенхимных клеток, а также постепенным вызреванием элементов радужной оболочки, смещающейся назад [6, 11, 14, 37, 89].
К пятому месяцу угол передней камеры округляется, и такая его форма сохраняется до седьмого месяца.
С третьего месяца отмечается постепенное углубление угла, которое продолжается даже после рождения на протяжении длительного времени (до 4 лет). На 7-м месяце эмбриональ-
Развитие передней камеры и дренажной системы
579
ного развития самая глубокая часть угла приближается к уровню расположения шлеммова канала, а на момент рождения лежит на уровне склеральной шпоры.
Таким образом, угол передней камеры новорожденного отличается от угла взрослых тем, что большая часть увеальной части трабекуляр-ной сети лежит впереди ресничной мышцы и перед склеральной шпорой.
Развитие трабекулярной сети (рис. 5.3.3). К четвертому месяцу эмбрионального развития зачаток трабекулярной сети представляет собой структуру треугольной или клиновидной формы, состоящую из недифференцированных эктомезенхимных клеток. Передняя часть этой массы клеток лежит между эндотелием роговой оболочки и наиболее глубоко лежащей стромой [6, 14, 49, 58, 63]. На этой стадии эндотелий роговицы закрывает большую часть передней поверхности трабекулярной сети, таким образом очерчивая границу передней камеры глаза. Сзади нет никакой границы между эктомезен-химными клетками, превращающимися в последующем в трабекулярную сеть, и клетками будущей ресничной мышцы.
Эктомезенхимные клетки трабекулярной сети располагаются первоначально свободно. Их цитоплазма содержит расширенные цистерны эндоплазматической сети, рибосомы и лизосо-мы. Между ними видны многочисленные кол-лагеновые волокна, а также немиелинизиро-ванные нервы.
Между четвертым и восьмым месяцами развития эктомезенхимные клетки удлиняются и продолжают секретировать коллагеновые волокна, эластическую ткань и основное вещест-
во [197]. Следующие изменения строения будущей трабекулярной сети наступают после развития склеральной шпоры. Это происходит между 22-й и 24-й неделями эмбрионального развития. Именно в это время мезенхимные клетки разделяются на наружную корнеоскле-ральную часть, ориентированную продольно, и внутреннюю увеальную часть, сохраняющую сетеподобное строение. Трабекулы корнеоскле-ральной части уже состоят из стержня коллаге-новых и эластических волокон, покрытых клетками, лежащими на базальной мембране. Благодаря наличию многочисленных цитоплазмати-ческих отростков клетки контактируют между собой.
Увеальная часть трабекулярного аппарата, в отличие от корнеосклеральной, пока еще состоит из недифференцированных эктомезенхимных клеток.
Необходимо отметить, что окончательная дифференциация и четкая ориентация трабекул зависит от прилагаемой к ним механической силы натяжения, направленной в определенном направлении. Под действием этой силы кор-неосклеральная и глубже расположенная увеальная части трабекулярного аппарата располагаются циркулярно, а внутренняя часть увеаль-ных трабекул ориентирована меридианально. Такое расположение позволяет создать более жесткую и одновременно упругую систему [262].
Между 28-й и 30-й неделями эмбрионального развития корнеосклеральные тяжи удлиняются, а между трабекулярными клетками появляются межклеточные контакты. К девятому месяцу между трабекулами увеальной части уже видны широкие межтрабекулярные про-
|
Дата добавления: 2018-10-18; Мы поможем в написании ваших работ!; просмотров: 169 | Нарушение авторских прав Поиск на сайте: Лучшие изречения:  | 4069 - | 4069 -  |