




В историческом плане вопрос об эволюции генов является важнейшим, поскольку эволюция генов связана с истоками жизни вообще и ее совершенствованием в частности. Поскольку выявлена изначальная роль в происхождении жизни РНК, то предполагают, что начало эволюции генов датируется 3,5-4 млрд лет назад, когда сформировались первые молекулы РНК, которые каким-то образом детерминировали синтез белков, т. е. были первыми хранителями генетической информации. Однако когда выявилась необходимость в повышении эффективности синтеза белков, способность кодирования генетической информации перешла к ДНК, которая стала главным хранителем генетической информации. Что касается РНК, то она оказалась между ДНК и белком, став «переносчиком» информации. Конечно, эта гипотеза не имеет доказательств. Тем не менее многие далее считают, что появление ДНК связано с усложнением структуры клеток и, следовательно, необходимостью кодирования большого количества информации по сравнению с РНК. Другими словами, с началом участия ДНК в хранении генетической информации стал развиваться генетический код.
В последнее время большое внимание приобрела гипотеза, в соответствии с которой источником новых генов является рекомбинация эксонов, а также транспозоны, поступающие в геномы организмов.
Особый интерес в эволюционном плане представляет ДНК, которая не транскрибируется (эгоистическая ДНК). Казалось, должны быть какие-то факторы контрселекции, которые обеспечивают поддержание этой ДНК в клетках. Между тем такие факторы неизвестны. Тем не менее очень популярно предположение, что эгоистическая ДНК тоже является источником образования новых генов.
В обсуждении направления эволюции геномов известно два объяснения. Одни ученые предполагают, что увеличение геномов клеток в процессе эволюции организмов шло путем включения в ядерные структуры дополнительных копий генов, в то время как другие считают, что в эволюции шла дупликация уже образованных генов с последующей их дивергенцией. Доказательства включения генов в геномы отсутствуют, тогда как предположение о дупликации и дивергенции генов имеет существенные обоснования, причем эти обоснования исходят из данных о том, что многочисленные семейства белков кодируются наборами родственнных генов.
Установлено, например, что в кодировании яичного альбумина цыплят участвует несколько генов. Лейкоцитарный интерферон человека кодируется девятью неаллельными генами, а 8-кристаллин цыплят — двумя генами.
Белок актин у насекомых и морских ежей контролируется тоже несколькими генами, причем для каждого актина в разных сократительных клетках существует свой ген. Белки хориона контролируются тоже несколькими генами, причем объединенными в сложный кластер. Идентифицировано несколько генов, кодирующих родопсины, обеспечивающие восприятие разных цветов. Наконец, иммуноглобулины также кодируются многими генами, например, у мышей тремя генами. Известны и другие примеры. Поэтому можно сказать, что синтез лишь нескольких белков контролируется одиночными генами. Например, одним геном кодируется синтез инсулина человека и кур. В подтверждение предположения о дупликации и дивергенции генов и, следовательно, о механизмах увеличения геномов наиболее показательны данные об эволюции мультисемейства генов, кодирующих гемоглобины.
У морских червей, некоторых насекомых и рыб размеры глобиновой молекулы, переносящей кислород, составляют всего лишь 150 аминокислотных остатков. У человека каждая молекула гемоглобина состоит из двух a-полипептидных цепей и двух b-полипептидных цепей.
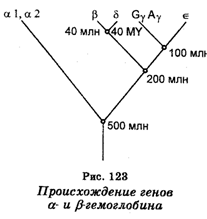 Синтез гемоглобинов кодируется двумя несцепленными кластерами генов. Кластер глобиновых генов локализован на 16 хромосоме и содержит два эмбриональных, а также два почти идентичных фетальных глобиновых гена. Второй кластер, располагающийся на хромосоме II, кодирует b-глобины. Он состоит из b-глобинового гена, одного эмбрионального e-гена, двух фетальных генов, а также гена d-глобина.
Синтез гемоглобинов кодируется двумя несцепленными кластерами генов. Кластер глобиновых генов локализован на 16 хромосоме и содержит два эмбриональных, а также два почти идентичных фетальных глобиновых гена. Второй кластер, располагающийся на хромосоме II, кодирует b-глобины. Он состоит из b-глобинового гена, одного эмбрионального e-гена, двух фетальных генов, а также гена d-глобина.
Исходя из данных о генетическом контроле глобинов и данных секвенирования глобиновых генов, предполагают, что самая первая дупликация гена, контролирующего синтез белка, являющегося предшественником гемоглобина, произошла 1 млрд лет назад, дав начало паре генов. Один ген из этой пары в ходе эволюции стал геном, контролирующим синтез миоглобина, другой развился в ген, контролирующий синтез гемоглобина. В последующем (около 500 млн лет назад) ген гемоглобина вновь подвергся дупликации, в результате чего возникли гены, контролирующие синтез гемоглобиновых цепей a и b. Через некоторое время дальнейшей дупликации подвергся ген b-цепи, что дало начало гену гемоглобина, синтезируемого только в эмбриональном периоде. Далее ген эмбрионального гемоглобина подвергся тоже дупликации, что дало начало уже гемоглоби-нам d и b. Еще одна дупликация гена b-цепи дала ген, кодирующий синтез e-глобина (рис. 123). Таким образом, в результате серии тандемных дупликации исходный глобиновый ген стал на одной из стадий развития кластерами генов a- и b-глобинов.
Эволюция геномов ускоряется транспозируемыми элементами, которые обладают способностью наращивать геномы. Наконец, увеличению геномов способствует генетическая рекомбинация.
Один из принципиальных вопросов связан с познанием механизмов, предупреждающих гены от дупликации в результате транскрипционной активации генов-«сосед ей». В недавнее время на примере D. melanogaster показано существование пограничных сегментов (последовательностей) ДНК, функционально изолирующих соседние гены. Длина последовательностей-изоляторов составляет около 340 пар оснований. Не исключено, что такие последовательности существуют и у других организмов.
Вопросы для обсуждения
1. На чем основаны представления о том, что генетическим материалом являются нуклеиновые кислоты? Каково значение генной инженерии в перечне доказательств генетической роли ДНК?
2. Существует ли связь между размерами генома (в количестве нукле-отидных пар) и видовой принадлежностью организмов? Приведите примеры в обоснование вашей точки зрения.
3. Что вы знаете о путях увеличения генома клеток в процессе развития организмов от низших форм к высшим?
4. Определите в сантиметрах общую длину ДНК в клетках человека.
5. ДНК стабильна при рН11, но РНК деградирует до нуклеотидов при щелочной реакции. Используйте учебники по биохимии и объясните причину этого явления.
6. Если субъединицы b и b' РНК-полимеразы составляют 0,005 доли от массы общего белка в клетках Е. coli, то сколько будет в клетке молекул РНК-полимеразы при условии, что каждая субъединица b и b' представляет цельную молекулу этого фермента?
7. Почему мочевина денатурирует РНК?
8. Состав оснований (фракция Г+Ц) двухцепочечной молекулы ДНК отражается на показателях плавучей плотности в хлориде цезия и температуры плавления (Тп), при которой половина молекул «плавится» на отдельные цепи. Было найдено, что плавучая плотность равна 1,660 + (0,098´фракция Г+Ц), фракция Г+Ц = 2,44 (Тп — 69,3), причем Tп определена в стандартном солевой растворе. Плавучая плотность ДНК крысы составляет 1,702, D. melanogaster — 1,698 и дрожжей — 1,699. Определите фракцию Г+Ц и температуру плавления для ДНК каждого вида.
9. Каково значение митохондриальной ДНК человека?
10. Что представляют собой транспозируемые генетические элементы? Как их классифицируют?
11. Что представляют собой плазмиды?
12. Что представляют собой повторяющиеся последовательности ДНК и как часто они повторяются в геноме человека?
13. В чем заключается полуконсервативный способ репликации ДНК и каково биологическое значение такого способа репликации?
14. Какова роль ферментов в репликации ДНК?
15. Есть ли разница между репликацией ДНК и репликацией хромосом?
16. Что такое нуклеосома и каковы ее размеры? Какова роль белков в упаковке ДНК в хромосомы?
17. Вычислите число нуклеотидных пар в 1 мегадальтоне двухцепочечной ДНК.
18. Как вы думаете, сколько генов имеется в одной клетке человека при условии, что длина одного гена составляет около 500 пар нуклеотидов?
19. По данным кислотного гидролиза препарат ДНК, выделенной из клеток мертворожденного плода человека, характеризовался следующим составом (в %): аденин — 25, тимин — 32, гуанин — 22, цитозин — 21. Каким образом можно объяснить этот необычный результат исследования, руководствуясь данными о структуре ДНК?
20. Какой будет длина и суммарная масса ДНК, если в ней объединить молекулы ДНК из всех клеток новорожденного ребенка, организм которого состоит из 2,6´1012 клеток?
21. Что представляют собой ферменты Оказаки и какова их роль в репликации ДНК?
22. Можете ли вы назвать экспериментальные данные, подтверждающие антипараллельную ориентацию цепей в молекуле ДНК?
23. После денатурации ДНК ренатурировали, позволив регибридиза-цию цепей до 1% их последовательностей. Затем эту ДНК подвергли обработке нуклеазой I до полного переваривания молекул, после чего ее «разогнали» путем электрофореза в агарозном геле. Каковы результаты электрофореза?
24. Каким образом можно определить генетическую локализацию IS1-последовательностей в геноме Е. coli?
25. Что собой представляют прямые и обратные мутации и каковы частоты этих мутаций по сравнению с супрессорными мутациями?
26. В чем заключаются молекулярные механизмы генных мутаций?
27. Опишите механизмы действия физических и химических мутагенов.
28. Могут ли восстанавливаться повреждения ДНК? Если да, то с помощью каких механизмов и какое значение это явление имеет в жизни организмов?
29. Можете ли вы объяснить расщепление ДНК в процессе ее восстановления с 5'-стороны от тиминового димера, но не с 3'-стороны?
Литература
Альберте В., Брей Д., Льюис Дж., Ряфф М., Роберте К., Уотсон Дж. Молекулярная биология клетки. М.: Мир. 1994. Т. 1. 815 стр.
Пехов А. П. Основы плазмидологии. М.: РУДН. 1998. 231 стр.
Karcher S. J. Molecular Biology. Academic Press. 1995. 273 pp.
Schleif R. Genetics and Molecular Biology. The Johns Hopldns University Press. 1983. 698 pp.
Глава XI
ЗАКОНОМЕРНОСТИ ПЕРЕДАЧИ
ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ.

