




(Богданов А.А., 1998)
| Тип клеток | Длина теломерной ДНК, тыс. п.н. | Теломеразная активность |
| Половые Соматические Раковые | 15-20 10-12 при рождении, уменьшается с возрастом 4-6, 10-15 | Высокая Отсутствует Присутствует в 80% случаев |
Транскрипция
Транскрипцией называют биосинтез РНК на матрице ДНК. Это первая стадия реализации генетической информации, в процессе которой определенные участки нуклеотидной последовательности ДНК «переписываются» в комплементарные одноцепочечные молекулы РНК. В результате транскрипции образуются мРНК, кодирующие аминокислотные последовательности белков, тРНК, рРНК и другие виды РНК, выполняющие структурные, регуляторные и каталитические функции.
Принципы транскрипции
Транскрипция во многом сходна с репликацией. В основе механизма копирования при транскрипции лежит фундаментальный принцип комплементарности азотистых оснований полинуклеотидных цепей ДНК и РНК. Как и при синтезе ДНК, субстратами синтеза РНК являются рибонуклеозид три фосфаты. В ходе включения в строящуюся цепь они теряют пирофосфатные остатки, что обеспечивает процесс энергией. Еще одно сходство с синтезом ДНК состоит в направлении роста строящейся цепи: 5′®3′, т. е. очередные нуклеотиды присоединяются к 3′-концу. Как и при всех матричных синтезах, строящаяся цепь антипараллельна матричной цепи ДНК.
Однако имеются и принципиальные отличия от синтеза ДНК. Транскрипция – ассиметричный процесс, в качестве матрицы используется лишь одна молекула ДНК, получившая название матричной или значащей. Синтез РНК не требует для своего начала никакой затравки. Процессу транскрипции подвергается единовременно не вся молекула ДНК, а только ее определенные участки – транскриптоны. Они ограничены двумя последовательностями, которые называются промотором (зона начала транскрипции) и терминатором (зона остановки транскрипции). Транскриптоны бактерий (опероны) обычно включают несколько структурных генов (цистронов). Поэтому синтезируемая на оперонах бактерий мРНК является полицистроновой и может быть использована для синтеза нескольких белков, в отличие от моноцистроновых мРНК эукариот, служащих матрицами для синтеза одного определенного белка.
РНК-полимеразы
Процесс транскрипции осуществляется с участием ферментов – РНК-полимераз, а также большой группы белков – регуляторов транскрипции.
У прокариот молекула РНК-полимеразы состоит из двух компонентов:
· минимальной РНК-полимеразы, или «кор»-фермента (от англ. core – серцевина), содержащей все каталитические центры, участвующие в синтезе РНК;
· σ-субъединицы, необходимой для правильного присоединения фермента к промотору и отделяющейся от РНК-полимеразы после начала синтеза РНК.
Комплекс минимального (сore -) фермента и σ-субъединицы называется холоферментом. C ore -фермент имеет одинаковое сродство к любой последовательности нуклеотидов (рис. 29а). Только полный (holo -) фермент обладает высоким сродством к промотору, сродство к остальным случайным последовательностям ДНК у него снижено в 10 тыс. раз
(рис. 29б).

Рис. 29. Инициация транскрипции ДНК бактериальной РНК-полимеразой
Как только произошла инициация транскрипции, σ-фактор отделяется. Элонгация (продолжение синтеза РНК) и терминация (остановка транскрипции) осуществляются core -ферментом. Минимальная РНК-полимераза состоит из четырех субъединиц: двух идентичных α-субъединиц и неидентичных β- и β′-субъединиц.
В ядрах эукариот обнаружены три специализированные формы РНК-полимеразы:
· РНК-полимераза I синтезирует рибосомные РНК (28S и 18S);
· РНК-полимераза II – матричные РНК;
· РНК-полимераза III – транспортные и низкомолекулярные рибосомные РНК (5S).
Этапы транскрипции
1. Инициация – первый этап транскрипции, в ходе которого происходит связывание РНК-полимеразы с промотором и образование первой межнуклеотидной связи.
У бактерий холофермент РНК-полимераза непосредственно узнает определенные последовательности нуклеотидных пар в составе промотора: последовательность 5′-ТАТААТ-3′ (расположена на расстоянии 10 нуклеотидов от точки начала транскрипции и называется боксом Прибнова) и последовательность 5′-ТТГАЦА-3′ (удалена от точки начала транскрипции на 35 нуклеотидов). В некоторых оперонах, например в лактозном, необходимо предварительное взаимодействие с промотором дополнительного белка (САР изменяет структуру промотора, резко повышая его сродство к РНК-полимеразе).
РНК-полимеразы эукариот не способны самостоятельно связываться с промоторами транскрибируемых генов. В присоединении к транскриптонам РНК-полимераз принимают участие общие факторы транскрипции (TF). Они отличаются от σ-факторов прокариот тем, что могут связываться с ДНК независимо от РНК-полимеразы. Полимеразы I, II и III требуют присутствия разных факторов транскрипции, обозначаемых TF I, TF II и TF III соответственно. Промоторы эукариот устроены более сложно, чем прокариотические, и состоят из нескольких элементов. Из низ самым близким к точке начала транскрипции является ТАТА-домен, называемый также доменом Хогнесса. Затем следуют домены ЦААТ и ГЦ. Промоторы эукариот могут содержать различные комбинации этих элементов, но ни один из них не встречается во всех промоторах. Домен ЦААТ играет существенную роль в инициации транскрипции, ТАТА и ГЦ, по-видимому, выполняют вспомогательные функции.
Связавшись с промотором, РНК-полимераза вызывает локальную денатурацию ДНК, т. е. разделение цепей ДНК на протяжении примерно 15 нуклеотидных пар. Образуется транскрипционный «глазок». Первым в строящуюся цепь РНК включается пуриновый нуклеотид – АТФ или ГТФ, при этом все три его фосфатных остатка сохраняются. После образования первой фосфодиэфирной связи σ-фактор у бактерий теряет связь с ферментом, и оставшийся core -фермент начинает перемещаться по ДНК. РНК-полимераза эукариот после инициации транскрипции также теряет связь с транскрипционными факторами и перемещается по ДНК самостоятельно.
2. Элонгация – последовательное удлинение растущей цепи РНК. Перемещаясь вдоль двойной спирали ДНК, РНК-полимераза непрерывно раскручивает спираль впереди того участка, где происходит синтез РНК. На короткое время образуется так называемый открытый комплекс, внутри которого возникает РНК-ДНК-спираль длиной около 20 нуклеотидов
(рис. 30). Затем фермент (с помощью специального сайта) вновь закручивает

Рис. 30. Элонгация транскрипции
ДНК позади участка полимеризации. РНК-транскрипт выводится из комплекса через особый канал, свойственный РНК-полимеразе.
Скорость синтеза РНК у бактерий составляет около 30 нуклеотидов в секунду, однако она не постоянна и может несколько снижаться. Такие периоды называют паузами транскрипции.
Показано, что еще до образования гибрида РНК-ДНК РНК-полимераза переводит ДНК из В-формы в А-форму. В ней плоскости азотистых оснований не перпендикулярны оси спирали, а наклонены на 200 к перпендикуляру. Вероятно, это облегчает разъединение двух соседних азотистых оснований в цепи ДНК. Параметры РНК-ДНК-спирали также практически полностью идентичны характеристикам А-формы ДНК.
3. Терминация (окончание транскрипции) определяется особой нуклеотидной последовательностью ДНК, расположенной в зоне терминатора оперона. В бактериальных оперонах выделяют два типа терминаторов:
· ρ(ро) -независимые терминаторы (I типа);
· ρ -зависимые терминаторы (II типа).
 ρ-независимые терминаторы состоят из последовательностей, представляющих собой инвертированный повтор – палиндром (рис. 31), и рас-
ρ-независимые терминаторы состоят из последовательностей, представляющих собой инвертированный повтор – палиндром (рис. 31), и рас-
Рис. 31. ρ -независимая терминация транскрипции у бактерий
полагаются за 16-20 нуклеотидных пар от точки терминации. Палиндромы (последовательности, которые читаются одинаково слева направо и справа налево) ρ- независимых терминаторов содержат большое количество Г-Ц-повторов. За этим участком на матричной цепи расположена олиго(А)-последовательность (4-8 адениловых нуклеотидов подряд). Транскрипция в области палиндрома приводит к тому, что в получившемся РНК-транскрипте быстро образуется устойчивый элемент вторичной структуры – «шпилька» – спирализованная область, содержащая комплементарные
Г-Ц-пары. «Шпилька» нарушает прочность связи ДНК-РНК в открытом комплексе. Кроме этого транскрипция олиго(А)-последовательности в матричной цепи ведет к образованию участка ДНК-РНК-гибрида, составленного из непрочных А-У пар, что также способствует разрушению контакта между ДНК и РНК.
ρ-зависимые терминаторы. Одним из факторов транскрипции прокариот является белок ρ. ρ -фактор – это имеющий четвертичную структуру белок, обладающий АТФ-азной активностью. Он способен связываться с 5'-концом синтезируемой РНК длиной около 50 нуклеотидов. ρ -фактор движется по РНК с такой же скоростью, с которой РНК-полимераза движется по ДНК. Вследствие того что в терминаторе много Г-Ц-пар (с тремя водородными связями), РНК-полимераза в области терминатора замедляет ход, ρ -фактор ее догоняет, изменяет конформацию фермента, и синтез РНК прекращается (рис. 32).
На терминаторах обоих типов происходят три ключевых события:
· останавливается синтез РНК;
· цепь РНК освобождается от ДНК;
· РНК-полимераза освобождается от ДНК.
Процессинг РНК
Первичные РНК (предшественники РНК, гетерогенные ядерные РНК), образующиеся в результате транскрипции, в большинстве случаев представляют собой функционально неактивные молекулы. Поэтому сразу после транскрипции они претерпевают ряд модификаций и превращаются в зрелые РНК. Созревание первичных транскриптов называется процессингом.
 Рис. 32. ρ- зависимая терминация транскрипции у бактерий
Рис. 32. ρ- зависимая терминация транскрипции у бактерий
Для бактериальных клеток процессинг предшественников мРНК не характерен и необходим только при образовании зрелых молекул рРНК и тРНК.
Процессинг рнк у эукариот представляет собой достаточно сложный и тонко организованный процесс, непосредственно влияющий на регуляцию экспрессии генетического материала. Наиболее детально изучен процессинг мРНК эукариот, который включает:
· сплайсинг – вырезание из пре-мРНК некодирующих областей (интронов) и сшивание кодирующих структуру белка участков (экзонов);
· кэпирование – образование на 5′-конце мРНК особой структуры – кэпа – происходит вскоре после начала синтеза мРНК и осуществляется с участием ГТФ;
· полиаденилирование – образование на 3′-конце поли(А)-фрагмента, содержащего около 200 адениловых нуклеотидов (рис. 33).

Рис. 33. Процессинг мРНК
Механизм сплайсинга
В сплайсинге пре-мРНК эукариот принимает участие ряд белков, а также РНК особого вида – малые ядерные РНК (мяРНК). Различные мяРНК по принципу комплементарности связываются с пограничными участками интронов РНК. Для этого взаимодействия существенны определенные последовательности нуклеотидов в начале и конце интронов: так, интроны всегда начинаются с Г-У, а заканчиваются дуплетом А-Г. Малые ядерные РНК образуют комплекс с ферментами, катализирующими сплайсинг – сплайосому.
Первый разрыв пре-РНК происходит в области 5′-конца интрона, который связывается с одним из нуклеотидов в средней части того же интрона (рис. 34). Это приводит к образованию кольцевой (или, точнее, лассоподобной) структуры. Первая мяРНК диссоциирует, а ферментный комплекс перемещается к другой мяРНК, маркирующей 3′-конец интрона. Здесь происходит второй разрыв пре-РНК. Связь экзона 2 с интроном заменяется связью с экзоном 1.
Альтернативный сплайсинг
В ряде случаев возможно изменение хода сплайсинга и осуществление его по альтернативному варианту. В этом случае с одного гена считывается более одного типа мРНК. Альтернативный сплайсинг позволяет организму синтезировать разные по структуре и свойствам белки на базе одного гена. Такие гены кодируют семейства родственных белков, участвующих в мышечных сокращениях, формировании цитоскелета, нервных  волокон, пептидных гормонов и др.
волокон, пептидных гормонов и др.
Рис. 34. Вероятный механизм спайсинга:
Е – ферментный комплекс (с нуклеазной и лигазной активностью)
Альтернативный сплайсинг мРНК включает три основных механизма:
1. Использование разных промоторов. При наличии в гене альтернативных промоторов разные типы РНК могут синтезироваться с разных сайтов инициации транскрипции. Альтернативный промотор – сложный промотор, состоящий по крайней мере из двух независимо функционирующих частей, расположенных перед разными экзонами одного гена. В этом случае образуются транскрипты, имеющие разные по длине 5′-концы и разное количество экзонов.
2. Изменение сайта полиаденилирования первичного транскрипта. В результате изменяются размеры и структура 3′-концевого участка пре-мРНК.
3. Соединение экзонов в различных комбинациях. При этом часть экзонов может не включаться в сплайсинг. Например, если ген содержит всего шесть экзонов (с 1-го по 6-й), в одном типе мРНК они могут располагаться в порядке 1,2,3,4,5,6, в других РНК порядок может быть иным, например 4,5,6,1,2,3, или 2,5,6, или 1,3,5.
Альтернативный сплайсинг обеспечивает тонкую регуляцию работы генов у эукариот, дифференцировку тканей, определяет развитие различных признаков, детерминированных одним геном. У человека около 1/3 всех генов может кодировать более одного белка, т. е. разные белки кодируются разными сочетаниями экзонов одного и того же гена. Наличие альтернативного сплайсинга может объяснить тот факт, что количество белков в организме человека в несколько раз больше, чем число белок-кодирующих генов.
Трансляция
Трансляция – процесс перевода генетической информации с последовательности нуклеотидов мРНК в последовательность аминокислот в молекуле полипептида. Трансляция осуществляется согласно правилам генетического кода, который имеет следующие особенности:
1. Код – триплетный, т. е. одну аминокислоту определяет три нуклеотида.
2. Код – однозначный (специфичный): каждый кодон обозначает только одну аминокислоту.
3. Код – непрерывный, т. е. отсутствуют сигналы, показывающие конец одного кодона и начало следующего.
4. Код – вырожденный, т. е. одной аминокислоте может соответствовать более одного кодона. Только две аминокислоты – метионин и триптофан – имеют по одному кодону. Лейцину и серину соответствует 6 кодонов, глицину и аланину – по 4 и т. д. Если аминокислота кодируется несколькими кодонами, то в большинстве случаев они различаются по третьей букве, т. е. по нуклеотиду на 3′-конце. Таким образом, специфичность каждого кодона определяется главным образом его первыми двумя нуклеотидами.
5. Код не перекрывается, т. е. один нуклеотид не может одновременно входить в два соседних триплета.
6. Генетический код содержит триплеты, обозначающие начало и окончание синтеза белка. АУГ – инициирующий кодон (кодирует метионин). УАА, УАГ, УГА – терминирующие кодоны, которые не кодируют ни одну из известных аминокислот и сигнализируют об окончании синтеза белка.
7. Генетический код универсален, т. е. одинаков у животных, растений, многих бактерий.
Подготовительные стадии
Подготовительные стадии трансляции включают:
· активацию аминокислот;
· присоединение аминокислот к тРНК.
 Обе стадии осуществляются с помощью фермента – аминоацил-тРНК-синтетазы (АРС-азы, кодазы). Существует 20 видов таких ферментов – по числу аминокислот. В каждом случае фермент имеет два центра узнавания – для аминокислоты и тРНК (рис. 35).
Обе стадии осуществляются с помощью фермента – аминоацил-тРНК-синтетазы (АРС-азы, кодазы). Существует 20 видов таких ферментов – по числу аминокислот. В каждом случае фермент имеет два центра узнавания – для аминокислоты и тРНК (рис. 35).
Рис. 35. Связывание аминокислоты (фенилаланина – Phe) с тРНК
В активном центре фермента аминокислота связывается с АТФ, лишь затем переносится на тРНК. Образование макроэргической связи между аминокислотой и тРНК называется аминоацилированием, а образовавшийся комплекс – аминоацил-тРНК (аа-тРНК). Каждая тРНК может переносить к месту синтеза белка только одну из аминокислот. Для большей части аминокислот имеется несколько тРНК, которые называются изоакцепторными и обозначаются соответственно тРНК1Phe, тРНК2Phe и т. д.
Этапы трансляции
Собственно процесс трансляции включает три фазы:
· инициацию;
· элонгацию;
· терминацию.
Инициация трансляции – начало синтеза полипептидной цепи, заключается в сборке белоксинтезирующей системы (активной рибосомы).
Функциональные центры рибосом
Каждая рибосома состоит из двух субчастиц: большой и малой. Форма субчастиц, их контактирующих поверхностей, достаточно сложная (рис. 36). На контактирующих поверхностях большой и малой субчастиц в небольших углублениях находятся центры связывания всех компонентов белоксинтезирующей системы (мРНК, пептидил-тРНК, очередная аминоацил-тРНК), а также центры, катализирующие образование пептидной связи и постепенное перемещение рибосомы относительно мРНК.


Рис. 36. Модель рибосомы Escherichia coli (Васильев В.Д., Институт белка РАН):
слева – перекрывающаяся проекция: малая (30S) субчастица обращена к зрителю
и закрывает собой часть большой (50S) субчастицы; справа – боковая проекция:
к зрителю обращен боковой палочкообразный выступ большой (50S) субчастицы,
а малая (30S) субчастица расположена вверху
Функциональные центры рибосом (рис. 37):
1.  Центр связывания мРНК (М-центр). Образован участком 18S-рРНК, который комплементарен на протяжении 5-9 нуклеотидов 5′-нетранслируемому фрагменту мРНК. Расположен на малой субчастице рибосомы.
Центр связывания мРНК (М-центр). Образован участком 18S-рРНК, который комплементарен на протяжении 5-9 нуклеотидов 5′-нетранслируемому фрагменту мРНК. Расположен на малой субчастице рибосомы.
2. Пептидильный центр (П-центр). В начале процесса трансляции с пептидильным центром связывается инициирующая аа-тРНК. На последующих стадиях трансляции в пептидильном центре находится пептидил-тРНК, содержащая уже синтезированную часть пептидной цепи.
3. Аминоацильный центр (А-центр) – место связывания очередной аа-тРНК. Аминоацильный и пептидильный центры расположены как на большой, так и на малой субчастицах рибосомы.
4. Каталитический (пептидилтрансферазный) центр (К-центр). Катализирует перенос пептидила из состава пептидил-тРНК на поступившую в амино-ацильный центр очередную аа-тРНК. Расположен на большой субчастице рибосомы.
Инициация трансляции у прокариот начинается со связывания мРНК в области 5′-нетранслируемого участка с малой субъединицей рибосомы. Инициирующий кодон (АУГ) оказывается на уровне пептидильного центра будущей рибосомы. Далее за счет комплементарного взаимодействия с этим кодоном происходит связывание инициирующей аа-тРНК. У прокариот инициирующей аа-тРНК является формилметиониновая аа-тРНК – fМet-тРНКi fMet
(рис. 38). Блокирование аминогруппы метионина формильным остатком препятствует включению такой аминокислоты во внутренние участки цепи, но в то же время позволяет fМet-тРНКi fMet связываться с инициирующим кодоном мРНК (АУГ). Инициирующая аа-тРНК, взаимодействуя с пептидильным центром большой субъединицы, вызывает связывание последней.
У прокариот инициация осуществляется при участии трех специфических белков – факторов инициации (IF – I nitiation F actors). IF-3, присоединяясь к малой субчастице рибосомы, препятствует преждевременному связыванию большой субчастицы и, с другой стороны, способствует связыванию мРНК. IF-2 участвует в связывании инициирующей аа-тРНК. Вероятно, этот фактор образует комплекс с аа-тРНК еще вне рибосомы, причем в состав комплекса входит ГТФ. В результате образуется так называемый инициаторный комплекс, состоящий из малой субчастицы рибосомы, мРНК, инициаторной аминоацил-тРНК и факторов инициации (рис. 39). Большая субчастица при ассоциации с малой субчастицей вызывает гидролиз ГТФ (до ГДФ и Фн) и одновременно вытесняет все факторы инициации, включая IF-3. В итоге инициации трансляции образуется полная 70S (у прокариот) рибосома с пептидильнымучастком, занятым инициаторной формилметионил-тРНК, и со свободным аминоацильнымучастком.

Рис. 39. Инициация трансляции у прокариот
Элонгация трансляции – основной и самый продолжительный этап белкового синтеза, в ходе которого происходит удлинение полипептидной цепи за счет последовательного присоединения аминокислот. Начинается с момента образования первой пептидной связи и заканчивается после включения в полипептидную цепь последней аминокислоты.
Элонгация у бактерий осуществляется при участии трех белковых факторов (EF-Tu, EF-Ts, EF-G) и имеет циклический характер.
Цикл элонгации включает 3 стадии:
1. Связывание аа-тРНК с аминоацильным центром рибосомы. На этой стадии со свободным А-центром рибосомы связывается очередная аа-тРНК – та, чей антикодон комплементарен кодону мРНК, находящемуся в А-центре. Поступив в А-центр, аа-тРНК закрепляется в нем в комплексе с белковым фактором EF-Tu (EF – E longation F actor) и ГТФ. При участии фактора EF-Тu осуществляется гидролиз ГТФ до ГДФ и Фн, а выделяющаяся энергия расходуется на сближение двух аминокислотных остатков. Комплекс EF-Tu·ГДФ при этом покидает рибосому и регенерируется с участием фактора EF-Ts, так что фактор EF-Tu вновь оказывается связанным с молекулой ГТФ (рис. 40).
 Рис. 40. Этап элонгации в синтезе белка у прокариот
Рис. 40. Этап элонгации в синтезе белка у прокариот
2. Образование пептидной связи. В рибосоме после первой стадии цикла находятся пептидил-тРНК (в П-центре) и аа-тРНК (в А-центре). При этом их акцепторные петли и связанные с ними аминокислотные остатки располагаются в каталитическом (К-) центре. Последний и осуществляет пептидилтрансферазную реакцию: переносит пептидил (или инициирующую аминокислоту – формилметионин у прокариот) на аминокислоту поступившей аа-тРНК. Прежняя тРНК пептидила становится свободной (рис. 40).

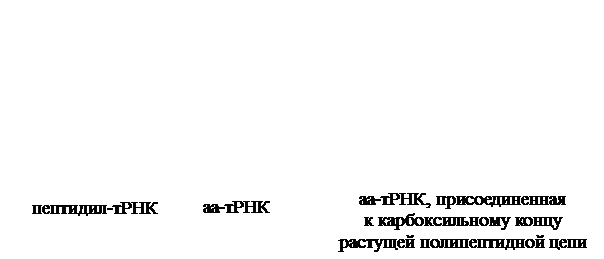 В ходе пептидилтрасферазной реакции карбоксильная группа пептидила образует пептидную связь с аминогруппой очередной аминокислоты (рис. 41). Таким образом, рост пептидной цепи при трансляции происходит в направлении от N- к С-концу.
В ходе пептидилтрасферазной реакции карбоксильная группа пептидила образует пептидную связь с аминогруппой очередной аминокислоты (рис. 41). Таким образом, рост пептидной цепи при трансляции происходит в направлении от N- к С-концу.
Рис. 41. Пептидилтрансферазная реакция
3. Транслокация – перемещение пептидил-тРНК из А-центра в П-центр в результате передвижения рибосомы по мРНК на один кодон. Свободная тРНК вытесняется из рибосомы, и одновременно освобождается А-центр, необходимый для связывания следующей аа-тРНК. Транслокация идет с участием белкового фактора EF-G (у бактерий) и сопровождается гидролизом одной молекулы ГТФ.
Таким образом, удлинение пептидной цепи на один аминокислотный остаток требует расхода двух молекул ГТФ (одна идет на связывание аа-тРНК, вторая – на траслокацию). Многократное повторение циклов элонгации приводит к включению в строящуюся пептидную цепь аминокислотных остатков в соответствии с последовательностью кодонов в мРНК.
Терминация трансляции. Сигналом об окончании трансляции служит появление в рибосоме одного из терминирующих кодонов мРНК: УАА, УАГ или УГА. С терминирующим кодоном, находящимся в А-центре, взаимодействуют особые белки – факторы терминации, или рилизинг-факторы (от англ. relеase – освобождать). У бактерий в терминации трансляции участвуют три белковых фактора: RF-1, RF-2 и RF-3. Фактор RF-1 узнает кодоны УАА и УАГ, а фактор RF-2 – кодоны УАА и УГА. Фактор RF-3 выполняет вспомогательную роль, стимулируя работу RF-1 и RF-2. При поступлении в рибосому одного из терминирующих кодонов с ним немедленно связывается соответствующий RF-фактор и тем самым блокирует присоединение аа-тРНК. Присоединение факторов терминации стимулирует гидролизную активность пептидилтрасферазного (каталитического) центра, в результате чего связь полипептида с тРНК гидролизуется. Синтезированный белок отделяется от рибосомы, одновременно отделяются тРНК и мРНК, а рибосома диссоцирует на субчастицы (рис. 42).
 В терминации трансляции принимает участие молекула ГТФ, которая, вероятно, служит аллостерическим регулятором активности белковых факторов терминации.
В терминации трансляции принимает участие молекула ГТФ, которая, вероятно, служит аллостерическим регулятором активности белковых факторов терминации.
Рис. 42. Терминация синтеза пептидной цепи у бактерий
Формирование пространственной структуры белков
После окончания трансляции процесс образования белков обычно еще не завершен. Для того чтобы белок приобрел присущие ему функциональные свойства, полипептидная цепь должна определенным образом свернуться в пространстве, сформировав функционально активную («нативную») структуру. Процесс формирования пространственной структуры белка называется фолдингом.
Стадии фолдинга
Пространственная (третичная) структура белка характеризуется сочетанием элементов вторичной структуры (α-спиралей, β-слоев), а также гибких участков полипептидной цепи, называемых петлями. Процесс сворачивания полипептидной цепи имеет ступенчатый характер (рис. 43) и включает:

Рис. 43. Стадии фолдинга
1) формирование элементов вторичной структуры;
2) специфическую ассоциацию некоторых элементов вторичной структуры с образованием супервторичной структуры: это могут быть сочетания нескольких α-спиралей, β-слоев или смешанные комплексы данных элементов;
3) промежуточную стадию, связанную с формированием основных элементов третичной структуры и образованием гидрофобного ядра молекулы. Молекула приобретает пространственную структуру, близкую к структуре нативного белка. Однако она еще не обладает присущей данному белку функциональной активностью. Это состояние, получившее название «расплавленная глобула», отличается от нативного меньшей степенью упорядоченности структуры (рис. 44);
4) образование нативной третичной структуры белка.
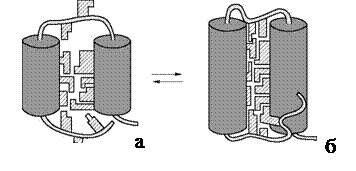
Рис. 44. Схема структуры «расплавленной глобулы» (а) и структуры
нативного белка (б): элементы вторичной структуры представлены
двумя спиральными участками (цилиндры); заштрихованные фигуры –
неполярные группы аминокислотных остатков
Факторы фолдинга опеспечивают оптимальные условия для реализации быстрого и эффективного образования нативной пространственной структуры. Выделяют две группы факторов:
1. Фолдазы – ферменты фолдинга (белки с каталитической активностью). Как и все ферменты, фолдазы необходимы лишь в каталитических количествах, т. е. в концентрациях, на порядки меньших, чем у «обслуживаемых» ими белков.
2. Молекулярные шапероны. Требуются в количествах, близких к стехиометрическим, т. е. сравнимых по величине с концентрацией сворачиваемых белков. Как и фолдазы, не входят в состав конечных продуктов фолдинга.
Часто построенная на рибосоме полипептидная цепь не может принять окончательную биологически активную (нативную) конформацию, пока не подвергнется процессингу, или ковалентной модификации. При этом может происходить отщепление ферментами инициирующих аминокислотных остатков, введение в определенные аминокислотные остатки фосфатных, метильных, карбоксильных, олигосахаридных или простетических групп. Например, гормоны поджелудочной железы инсулин и глюкагон синтезируются в виде неактивных предшественников. Сначала из них удаляются сигнальные пептиды, в результате чего образуются прогормоны. Затем уже в секреторных гранулах прогормоны превращаются в активные гормоны.
Новообразованные белки направляются в ту часть клетки, где они выполняют свои функции: некоторые поступают в клеточный цитозоль, другие − к различным клеточным органеллам, третьи секретируются из клетки, наконец, некоторые встраиваются в ту или иную клеточную мембрану, где работают в качестве ферментов или переносчиков.

