




Сопряжение анаболических и катаболических процессов
Обмен веществ, происходящий в живом организме, называют метаболизмом. Часть метаболизма, заключающаяся в процессах синтеза, называется анаболизмом, а соединения, участвующие в обмене веществ и подвергающиеся взаимному превращению, называются метаболитами. Его другая часть, связанная с процессами распада или деградации веществ, называется катаболизмом. Все процессы биосинтеза веществ нуждаются в энергии. Реакции, идущие с потреблением энергии, называют эндергоническими. Катаболитические процессы, как правило, протекают с выделением энергии и называются экзергоническими. Обычно энергия катаболитических процессов используется в анаболитических реакциях.
Ключевыми соединением в реакциях переноса энергии является АТФ. Говорят, что энергия катаболитических реакций запасается в форме АТФ и расходуется в анаболитических процессах при участии АТФ.
Процессы анаболизма и катаболизма являются сопряженными, т.е. связанными между собой, и в процессе сопряжения происходит передача энергии, запасенной в форме АТФ. АТФ - энергетическая валюта клетки.
Перенос энергии в живой природе
Все процессы жизнедеятельности сопровождаются переносом энергии. Живые существа по способу получения энергии делят на 2 категории:
- автотрофы - синтезируют сложные соединения из углекислого газа,
- - фотоавтотрофы - в качестве источника энергии используют солнечный свет,
- - хемоавтотрофы - получают энергию при окислении неорганических веществ,
- гетеротрофы - получают необходимую энергию за счет окисления сложных органических соединений.
Перенос энергии в биологических системах характеризуется
следующими особенностями:
1. Для биологических реакций характерны относительно небольшие изменения энергии. Если при расщеплении метаболита А до конечного продукта Р количество освобожденной энергии велико, то реакция в клетке обычно разделяется на несколько промежуточных стадий, при протекании которых выделяется незначительное количество энергии. С одной стороны, такая разбивка обеспечивает лучший выход продукта, а с другой - позволяет осуществлять ее регуляцию на промежуточных этапах.
2. Энергия, высвобождающаяся в экзергонических реакциях, запасается в виде АТФ (или других энергетически емких молекул). Все реакции анаболизма носят эндергонический характер, поэтому они должны быть сопряжены с экзергоническими реакциями. В качестве основного связующего звена выступает АТФ.
Например, химические связи в молекуле АТФ условно подразделяют на бедные энергией (при их разрыве выделяется менее 5 ккал/моль) и богатые энергией (при их разрыве выделяется более 5 ккал/моль). Богатые энергией связи называют макроэргическими.
АТФ: А-r-P-O~P-O~P
↑ ↑
7.3 7.6 ккал/моль
3. Все катаболитические реакции, имеющие место в живых организмах, в результате которых освобождается энергия при окислении субстратов, можно разбить на 3 группы:
а) Гидролитические реакции, в результате которых происходит превращение сложных молекул субстратов в относительно небольшое число метаболитов. Например, углеводы превращаются в моносахариды, липиды гидролизуются до спиртов и жирных кислот, белки гидролизуются до аминокислот. В результате таких превращений выделяется около 0.1 % от всей энергии, получаемой организмом.
б) Простые метаболиты подвергаются специфическим реакциям расщепления, в результате которых выделяется сравнительно небольшое количество энергии:
- гексозы путем гликолиза расщепляются до пировиноградной кислоты,
- жирные кислоты путем β-окисления превращаются до активной формы уксусной кислоты – ацетилкофермента А,
- аминокислоты путем дезаминирования пре5вращаются в оксикислоты.
В результате данных процессов окисления получаются ценные продукты: СН3СООН, СН3СО~СоА, кислоты (фумаровая, щавелево-уксусная), называемые центроболитами. Центроболиты могут быть далее подвергнуты дальнейшему окислению и получению энергии или же использоваться в анаболитических процессах в качестве строительного клеточного материала. На данном этапе выделяется около 1/3 всей энергии, получаемой при окислении субстрата.
в) Цикл реакций, в процессе которых метаболиты расщепляются до углекислого газа и воды - цикл Кребса (ЦТК) и Дыхательная цепь. Именно эти реакции обеспечивают в основном все энергетические потребности клетки. На данной стадии организм получает 2/3 всей энергии окисленного субстрата.
Процессы окисления в биологических системах подразделяются на аэробные и анаэробные.
Аэробное окисление - процесс, протекающий при достаточном количестве кислорода.
Анаэробное окисление - процесс, протекающий в условиях дефицита кислорода.
ФЕРМЕНТЫ
Общее понятие
Помимо того, что белки являются структурным компонентом клеток и тканей, наиболее важной является каталитическая активность белков.
В живых организмах протекает одновременно множество реакций легко и быстро благодаря присутствию специальных биокатализаторов ферментов. Например, гидролиз белка.
Катализаторы - вещества, меняющие скорость химических реакций, но не входящие в состав готовых продуктов.
Вещества, подвергающиеся превращениям в процессе ферментативных реакций, называют субстратами (s).
Ферменты значительно эффективнее обычных катализаторов. Это связано с тем, что они обладают высокой специфичностью по отношению к катализируемым реакциям и могут катализировать либо одну, либо небольшое число реакций. Непременным условием их действия является связывание с реагирующей молекулой (субстратом) и образование фермент-субстратного комплекса.
Действие катализаторов заключается в том, что они снижают энергию активации (Еа), необходимую для осуществления данной реакции, путем образования промежуточного комплекса фермент-субстрат.
сахароза → глюкоза + фруктоза
| Катализатор | Энергия активации, кал/моль |
| отсутствует | |
| НСl | |
| Сахараза |
Понижение энергии активации субстрата-сахарозы происходит из-за деформации молекулы субстрата, происходящей при образовании субстрат-ферментного комплекса. Деформация ослабляет внутримолекулярные связи и делает молекулу значительно более способной к определенной реакции.
Строение ферментов
Ферменты могут иметь различное строение: они могут состоять только из белка, обладающего каталитическими свойствами, или же из белка – апофермента и небелковой части - простетической группы.
Если простетическая группа, легко отделяется от белковой части молекулы фермента, ее называют коферментом. Коферменты непосредственно участвуют в реакции, катализируемой ферментом, а белковая часть молекулы – апофермент - определяет специфичность реакции на этапе связывания субстрата.
В молекуле каждого фермента выделяют активный центр (АЦ) – это та часть молекулы фермента, к которой присоединяется субстрат. На АЦ приходится малая часть объема молекулы фермента.
В составе АЦ фактически следует различать участок, ответственный за присоединение субстрат - центр связывания и каталитический центр, который воздействует на субстрат, заставляя его вступать в реакцию и изменяться. В каталитических центрах большей части известных ферментов обычно находят один или несколько остатков из числа следующих аминокислот (АК): серин (ser), цистеин (cys), лизин (lys), тирозин (tyr), гистидин (his), называемых функциональными группами, которые непременно сближены в пространстве, а в линейной последовательности полипептида могут располагаться достаточно далеко друг от друга.
Коферменты, как правило, выполняют функцию каталитического центра.
Химическая природа коферментов очень разнообразна.
В качестве коферментов чаще всего могут выступать соединения нуклеотидной природы: никотинамидадениндинуклеотид (НАД) и его фосфоргый эфир (НАДФ), АТФ, Коэнзим-А. У ряда ферментов коферментами являются флавопротеиды: флавинадениндинуклетотид (ФАД). Функцию коферментов могут выполнять витамины.
Например, НАД и ФАД катализируют отщепление от субстратов водорода.
Коферментами могут являться:
1) Тетрапиррольные структуры (например, группа ГЕМ в молекуле гемоглобина), содержащие ионы металлов переменной валентности (например, Fe2+/3+). Эти структуры участвуют в переносе электронов и входят в состав ферментов: цитохромы, каталаза, гемоглобин.
2) Ионы металлов (Меt+): Zn2+, Mg2+, Mn2+, Ca2+, Fe2+, Cu2+, Mo2+.
3) Производные водорастворимых витаминов:
В1(тиамин) - тиаминпирофосфат. Вит.В1 в виде тиаминпирофосфата является коферментом декарбоксилаз кетокислот и участвует в процессах декарбоксилирования.
РР (никотинамид) - НАД, НАДФ. является составной частью коферментов НАД, НАДФ, которые катализируют реакции биологического окисления, участвуют в процессах распада углеводов, синтеза ЖК, обмене аминокислот (АК).
В2 (рибофлавин) - ФАД. Эти ферменты, содержащие ФАД, катализируют процессы тканевого дыхания, синтеза и распада жирных кислот (ЖК). Вит.В2 регулирует важные процессы обмена веществ в организме.
В6 (пиридоксаль). В виде фосфорного эфира фосфопиридоксаль выступает в качестве кофермента декарбоксилаз, трансаминаз и участвует в процессах декарбоксилирования и переаминирования аминокислот.
Свойства ферментов
Каждый фермент характеризуется таким показателем, как активность. За единицу активности принимают количество фермента, которое в стандартных условиях катализирует превращение 1 мкМоля субстрата в 1 минуту.
Кроме того, в 1972 г. было предложено выражать ферментативную активность в каталах. Катал - это количество фермента, катализирующее превращение 1 Моля субстрата в 1 секунду.
Существует понятие удельной активности. Это активность фермента, отнесенная к его молекулярной массе, если таковая известна (Е/мг белка, Е/мл белка).
Свойства ферментов:
1. Термолабильность. Ферменты - термолабильные вещества, чувствительные к изменению температуры. Температура, при которой каталитическая активность максимальна - называют температурным оптимумом (Тopt). Тopt большинства ферментов животного происхождения лежит в интервале 40-50оС, а растительного - 50-60оС. Однако, встречаются ферменты с Тopt = 80оС. При повышении температуры выше Тopt фермент инактивируется, т.е. его активность снижается вплоть до исчезновения.
2. Влияние рН среды. На активность фермента оказывает влияние концентрация ионов водорода. Для каждого фермента существует свое оптимальное значение рН, при котором его активность максимальна. Отклонение значения рН от оптимального вызывает понижение активности фермента вплоть до его полной инактивации.
Фермент рНopt Функция
Пепсин 1.5-2.0 гидролиз белка
Трипсин 7.8-8.5 гидролиз белка
Липаза 4.7-5.0 гидролиз жиров
3. Специфичность. Она заключается в том, что каждый фермент может катализировать одну или несколько близких по своей природе реакций.

|
Еще в 1890 г. Эмиль Фишер предсказал, что «фермент должен подходить субстрату, как ключ к замку». Это означает, что фермент может катализировать превращение субстрата, если имеет место соответствие пространственных конфигураций формы молекулы субстрата активному центру фермента. Такое соответствие часто называют сродством.
Однако значительно позднее Кошланд высказал идею об индуцированном соответствии пространственных конфигураций активного центра фермента субстрату, что впоследствии подтвердилось. При связывании субстрата происходит изменение формы фермента. Активный центр фермента только после присоединения субстрата становится комплементарным ему по форме.
Специфичность ферментов условно подразделяют на:
Относительную специфичность - ею обладают ферменты, которые действуют на соединения, имеющие определенный тип связей. Например, протеиназы действуют на пептидную группу. Это ферменты широкого спектра действия.
Групповая специфичность - характерна для ферментов, которые действуют на субстраты, имеющие одинаковый тип связи и одну из функциональных группировок. Это могут быть: фосфомоноэстеразы, гидролизующие субстрат определенной структуры, например, от фосфатидной кислоты отщепляют фосфорную группировку (―РО3Н2).
Абсолютная специфичность - ею обладают ферменты, действующие только на один субстрат с вполне определенной структурой.
К ферментам, обладающим абсолютной специфичностью относятся уреаза, катализирующая превращение карбамида (мочевины) до углекислого газа и воды, ацетилхолинэстераза и др.
Стереохимическая специфичность - фермент действует только на определенный стереоизомер.
Активаторы и ингибиторы
Каталитическая функция ферментов зависит от влияния различных веществ, одни из которых повышают скорость реакций, а другие – понижают. В соответствии с воздействием на фермент все вещества можно разделить на активаторы и ингибиторы.
Активаторы – вещества, повышающие активность ферментов. Они способны защищать ферменты от агрессивных химических воздействий. В ряде случаев, например, свободный цистеин защищает сульфгидрильные (SH-) группы цистеиновых остатков фермента от окисления. Mg2+ - активирует реакции с участием АДФ и АТФ.
Ингибиторы – вещества, угнетающие действие ферментов. Механизм их действия состоит в том что ингибитор вступает в соединение с ферментом, образуя неактивный ингибитор-фермент комплекс вместо комплекса фермент-субстрат, что приводит к блокированию фермента.
Ингибирование может быть неспецифическим и специфическим.
При неспецифическом ингибировании наблюдается действие солей тяжелых металлов (Pb2+; Hg2+; Cd2+), тонина; CCl3COOH на белковые молекулы, которые образуют с белками нерастворимые осадки, вызывая их осаждение. Его могут вызывать также концентрированные кислоты и щелочи, органические растворители.
Действие специфических ингибиторов основано на специфическом связывании с определенными группами в активном центре фермента. Например, СО (окись углерода) специфически ингибирует ряд окислительных ферментов, соединенных в активном центре Fe или Cu. Они вступают во взаимодействие с металлами, блокируют активный центр.
Ингибирование может быть обратимым и необратимым.
При необратимом ингибировании ингибитор ковалентно соединяется с ферментом или связывается очень прочно. Примером может служить действие нервно-паралитических газов на ацетилхолинэстеразу, играющую важную роль в пердаче нервных импульсов. При этом ингибитор приводит к необратимому изменению структуры фермента и его инактивации, являясь денатурирующим агентом. Необратимые ингибиторы - это сильные кислоты, щелочи, спирт, цианиды и т.д.
Примером обратимого ингибирования служит конкурентное ингибирование. В качестве конкурентных ингибиторов могут выступать соединения близкие по структуре к субстрату, обладающие иногда большим сродством к ферменту, чем субстрат. Они занимают место в активном центре фермента, блокируя его. Этот тип ингибирования является обратимым. Чем выше концентрация субстрата, тем меньше сказывается действие ингибитора. Например, малоновая кислота COOH-CH2-COOH – конкурентный ингибитор сукцинатдегидрогеназы, субстратом которой является янтарная кислота (сукцинат) COOH- CH2 -CH2-COOH.
Частным случаем конкурентного ингибирования является субстратное ингибирование. Оно наблюдается при слишком высоких концентрациях субстрата. В этом случае активный центр фермента блокируется за счет того, что одновременно несколько молекул пытаются с ним связаться.
Неконкурентное ингибирование. В этом случае ингибитор соединяется с ферментом не по активному центру, при этом меняется конфигурация всей молекулы фермента, в том числе и активного центра, ингибируя его (снижая его активность). Например, HCN действует на железосодержащие ферменты, осуществляющие перенос электронов при окислительно-восстановительных реакциях. Тяжелые металлы действуют путем присоединения по тиоловым группам ферментов. Их действие не снимается добавлением субстрата.
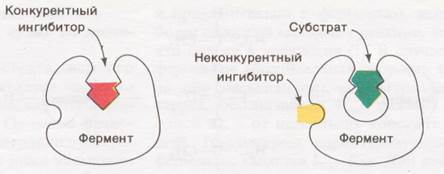
Часто активаторы и ингибиторы называют эффекторами.
Эффекторы – химические соединения, влияющие на ход ферментативных реакций. Они могут быть активаторами («+» эффекторами) или ингибиторами («-» эффекторами).
Классификация ферментов
Официальная классификация ферментов была принята в 1961 г. В соответствии с этой классификацией название фермента должно отражать тип катализируемой реакции.
Все ферменты подразделяют на 6 групп:
1. Оксидоредуктазы - катализируют реакции окисления и восстановления. Большая часть оксидоредуктаз имеет коферменты.
Оксидазы - ферменты, использующие для окисления субстрата молекулярный кислород.
Гидроксилазы - ферменты, преобразующие в присутствии О2
-СН3 → -СН2-ОН
-СН2- → -СН-ОН
| |
Дегидрогеназы - катализируют отщепление водорода от окисляемых субстратов.
2. Трансферазы - отрывают химическую группу от одного соединения, связывают ее, а затем присоединяют к другому соединению:
Трансаминазы - переносят аминогруппу с АК на кетокислоты.
Фосфокиназы (или просто киназы) - переносят остаток фосфорной кислоты на АДФ или от АТФ на субстрат.
3. Гидролазы - катализируют разрыв химической связи с присоединением молекулы воды.
Эстеразы - гидролизуют сложные эфиры с образованием кислоты и спирта.
Липазы - гидролизуют глицериды на глицерин и жирные кислоты.
Гликозидазы - гидролизуют гликозиды на сахарид и спирт.
4. Лиазы - катализируют разрыв связей С-С, C-N, C-O, C-S.
Декарбоксилазы - катализируют реакции декарбоксилирования:
R-CO-COOH → R-CHO + CO2.
Альдолазы - катализируют разрыв гексозофосфатов на 2 молекулы триозофосфатов.
5. Изомеразы - катаризируют различные процессы изомеризации.
Эпимеразы - вызывают взаимные переходы сахаров, например, галактоза ↔ глюкоза.
Мутазы - катализируют перенос химических групп с одной части молекулы на другую, например глюкозо-6 фосфатмутаза катализирует превращение глюкозо-6 фосфата в глюкозо-1 фосфат.
6. Лигазы - катализируют процессы конденсации двух сочетающихся молекул за счет энергии распада АТФ:
Аминоацил-тРНК-синтетазы - присоединяют АК к молекуле транспортной РНК. Это первый этап синтеза белка.

