




Тема 10.
Лекция 16. Биохимия печени
Химический состав печени.
Роль печени в обмене углеводов, липидов, белков.
Детоксикация различных веществ в печени.
| Краткое содержание | Конспект | ||||||||||||||||||||||||
| 1. Химический состав печени | |||||||||||||||||||||||||
| Печень функционирует как первичный регулятор содержания в крови веществ, поступающих в организм с пищей. Роль печени заключается в поддержании постоянства внутренней среды организма. Печень выполняет также крайне важную экскреторную функцию, теснейшим образом связанную с ее детоксикационной функцией. В организме нет путей обмена веществ, которые прямо или косвенно не контролировались бы печенью. Более половины сухого остатка печени приходится на долю белков, причем примерно 90% из них – на глобулины. | Химический состав печени млекопитающих
| ||||||||||||||||||||||||
| 2. Роль печени в обмене углеводов, липидов, белков | |||||||||||||||||||||||||
| Роль печени в углеводном обмене Основная роль печени в углеводном обмене заключается в обеспечении постоянства концентрации глюкозы в крови. Это достигается регуляцией между синтезом и распадом гликогена, депонируемого в печени. После приема пищи содержание глюкозы в воротной вене резко возрастает: в тех же пределах увеличивается и ее внутрипеченочная концентрация. Повышение концентрации глюкозы в печени вызывает существенное увеличение активности глюкокиназы и автоматически увели-чивает поглощение глюкозы печенью (образовавшийся глюкозо-6-фосфат либо затрачивается на синтез гликогена, либо расщепляется). В реакциях пентозофосфатного пути в печени образуется НАДФН, используемый для восстановительных реакций в процессах синтеза жирных кислот, холестерина и других стероидов. Наряду с утилизацией глюкозы в печени происходит и ее образование. Непосредственным источником глюкозы в печени служит гликоген. Кроме того, глюкоза в печени образуется также в процессе глюконеогенеза. В печени происходят превращения фруктозы и галактозы. Роль печени в липидном обмене Ферментные системы печени способны катализировать все реакции или значительное большинство реакций метаболизма липидов. Совокупность этих реакций лежит в основе таких процессов, как синтез высших жирных кислот, триглицеридов, фосфолипидов, холестерина и его эфиров, а также липолиз триглицеридов, окисление жирных кислот, образование ацетоновых (кетоновых) тел и т.д. Из печени кетоновые тела током крови доставляются в ткани и органы (мышцы, почки, мозг и др.), где они быстро окисляются при участии соответствующих ферментов Основным местом образования плазменных пре-β-липопротеинов (липопротеины очень низкой плотности – ЛПОНП) и α-липопротеинов (липопротеины высокой плотности – ЛПВП) является печень. Основной белок апопротеин В-100 (апо Б-100) липопротеинов синтезируется в рибосомах шероховатого эндоплазматического ретикулума гепатоцитов. В гладком эндоплазматическом ретикулуме, где синтезируются и липидные компоненты, происходит сборка ЛПОНП. В печени происходит интенсивный распад фосфолипидов, а также их синтез. Помимо глицерина и жирных кислот, которые входят в состав нейтральных жиров, для синтеза фосфолипидов необходимы неорганические фосфаты и азотистые соединения, в частности холин, для синтеза фосфатидхолина. Синтез фосфолипидов лимитируется количеством азотистых оснований, т.е. для синтеза фосфоглицеридов необходим либо холин, либо соединения, которые могут являться донорами метильных групп и участвовать в образовании холина (например, метионин). Такие соединения получили название липотропных веществ. Часть холестерина поступает в организм с пищей, но значительно большее количество его синтезируется в печени из ацетил-КоА. Биосинтез холестерина в печени подавляется экзогенным холестерином, т.е. получаемым с пищей. Часть синтезированного в печени холестерина выделяется из организма вместе с желчью, другая часть превращается в желчные кислоты и используется в других органах для синтеза стероидных гормонов и иных соединений. Роль печени в обмене белков Печень играет центральную роль в обмене белков. Она выполняет следующие основные функции: синтез специфических белков плазмы; образование мочевины и мочевой кислоты; синтез холина и креатина; трансаминирование и дезаминирование аминокислот, что весьма важно для взаимных превращений аминокислот, а также для процесса глюконеогенеза и образования кетоновых тел. Все альбумины * плазмы, 75–90% α-глобулинов и 50% β-глобулинов синтезируются гепатоцитами. Печень является единственным органом, где синтезируются такие важные для организма белки, как протромбин, фибриноген, проконвертин и проакцелерин. Патологический процесс в гепатоцитах резко снижает их синтетические возможности. В результате содержание альбумина в плазме крови резко падает, что может привести к снижению онкотического давления плазмы крови, развитию отеков, а затем асцита. В организме образование мочевины в основном происходит в печени. Синтез мочевины связан с затратой довольно значительного количества энергии (на образование 1 молекулы мочевины расходуется 3 молекулы АТФ). При заболевании печени, когда количество АТФ в гепатоцитах уменьшено, синтез мочевины нарушается. Большая часть мочевой кислоты также образуется в печени, где много фермента ксантиноксидазы, при участии которого оксипурины (гипоксантин и ксантин) превращаются в мочевую кислоту. Нельзя забывать о роли печени и в синтезе креатина. Имеются два источника креатина в организме. Существует экзогенный креатин, т.е. креатин пищевых продуктов (мясо, печень и др.), и эндогенный креатин, синтезирующийся в тканях. Синтез креатина происходит в основном в печени, откуда он с током крови поступает в мышечную ткань. Здесь креатин, фосфорилируясь, превращается в креатинфосфат, а из последнего образуется креатин. |
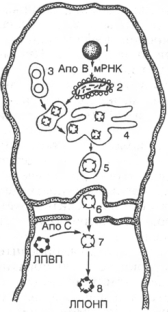 Образование липопротеинов очень низкой плотности (ЛПОНП) в печеночной клетке.
1 - ядро; 2 - шероховатый эндоплазматический ретикулум; 3 - гладкий эндоплазматический ретикулум, синтезированные в нем липиды и образовавшиеся н-ЛПОНП (незавершенные частицы); 4 - аппарат Гольджи; 5 - секреторная везикула с частицей н-ЛПОНП; 6 - частица с н-ЛПОНП в пространстве Диссе; 7 - перенос апопротеинов С с ЛПВП на н-ЛПОНП; 8 - частица нативных ЛПОНП.
Образование липопротеинов очень низкой плотности (ЛПОНП) в печеночной клетке.
1 - ядро; 2 - шероховатый эндоплазматический ретикулум; 3 - гладкий эндоплазматический ретикулум, синтезированные в нем липиды и образовавшиеся н-ЛПОНП (незавершенные частицы); 4 - аппарат Гольджи; 5 - секреторная везикула с частицей н-ЛПОНП; 6 - частица с н-ЛПОНП в пространстве Диссе; 7 - перенос апопротеинов С с ЛПВП на н-ЛПОНП; 8 - частица нативных ЛПОНП.
| ||||||||||||||||||||||||
| 3. Детоксикация различных веществ в печени | |||||||||||||||||||||||||
| Чужеродные вещества (ксенобиотики) в печени нередко превращаются в менее токсичные и даже индифферентные вещества. По-видимому, только в этом смысле можно говорить об «обезвреживании» их в печени. Происходит это путем окисления, восстановления, метилирования, ацетилирования и конъюгации с теми или иными веществами. Необходимо отметить, что в печени окисление, восстановление и гидролиз чужеродных соединений осуществляют в основном микросомальные ферменты. В результате гнилостных процессов, протекающих в кишечнике, из тирозина образуются фенол и крезол, а из триптофона – скатол и индол. Эти вещества всасываются и с током крови поступают в печень, где обезвреживаются путем образования парных соединений с серной или глюкуроновой кислотой. Обезвреживание фенола, крезола, скатола и индола в печени происходит в результате взаимодействия этих соединений не со свободными серной и глюкуроновой кислотами, а с их так называемыми активными формами: ФАФС и УДФГК. Примером обезвреживания токсичных продуктов в печени путем восстановления является превращение нитробензола в парааминофенол. Многие ароматические углеводы обезвреживаются путем окисления с образованием соответствующих карбоновых кислот. Печень принимает активное участие в инактивации различных гормонов. С током крови гормоны попадают в печень, при этом активность их в большинстве случаев резко снижается или полностью утрачивается. Так, стероидные гормоны, подвергаясь микросомальному окислению, инактивируются, превращаясь затем в соответствующие глюкурониды и сульфаты. Под влиянием аминооксидаз в печени происходит окисление катехоламинов и т.д. |
Метаболизм и выведение ксенобиотиков из организма
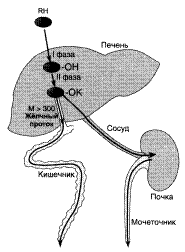 RH – ксенобиотик; К – группа, используемая при конъюгации (глутатион, глюкуронил и др.); М – молекулярная масса.
Система обезвреживания включает множество разнообразных ферментов, под действием которых практически любой ксенобиотик может быть модифицирован.
Микросомальные ферменты катализируют реакции С-гидроксилирования, N-гидроксилирования, О-, N-, S-дезалкилирования, окислительного дезаминирования, сульфоокисления и эпоксидирования.
RH – ксенобиотик; К – группа, используемая при конъюгации (глутатион, глюкуронил и др.); М – молекулярная масса.
Система обезвреживания включает множество разнообразных ферментов, под действием которых практически любой ксенобиотик может быть модифицирован.
Микросомальные ферменты катализируют реакции С-гидроксилирования, N-гидроксилирования, О-, N-, S-дезалкилирования, окислительного дезаминирования, сульфоокисления и эпоксидирования.
| ||||||||||||||||||||||||

