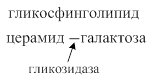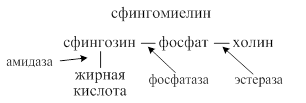Передача нервного импульса в синапсах осуществляется с помощью нейромедиаторов. Классическими нейромедиаторами являются ацетилхолин и норадреналин.
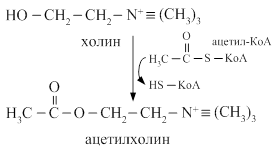
Ацетилхолин синтезируется из ацетил-КоА и холина при участии фермента ацетилхолинтрансферазы, накапливается в синаптических пузырьках, выделяется в синаптическую щель и взаимодействует с рецепторами постсинаптической мембраны. Ацетилхолин разрушается ферментом холинэстеразой.
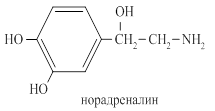
Норадреналин синтезируется из тирозина, разрушается ферментом моноаминоксидазой.
В качестве медиаторов могут выступать также ГАМК (гамма-аминомасляная кислота), серотонин, глицин.
Особенности метаболизма нервной ткани заключаются в следующем:
· наличие гематоэнцефалического барьера ограничивает проницаемость мозга для многих веществ;
· преобладают аэробные процессы;
· основным энергетическим субстратом является глюкоза.
Билет 22
Митохондриальная дых.цепь
Дыхательными цепями они называются в связи с тем, что очень часто внутримитохондриальное окисление называют тканевым дыханием. Последовательность компонентов цепи переноса электронов во внутренней мембране митохондрий определяется величиной их окислительно-восстановительного потенциала.
В начале ЦПЭ находятся вещества с наиболее отрицательным окислительно-восстановительным потенциалом. Эти вещества легче отдают электроны.. В митохондриях принято различать короткую и длинную дыхательные цепи.
Длинная дыхательная цепь
Длинная дыхательная цепь включает в себя окисление, начинающееся в матриксе митохондрий при участии НАД (НАДФ)- дегидрогеназ. В длинной цепи окисляются, например, изолимонная кислота, яблочная кислота, жирные кислоты, молочная кислота.
В матриксе происходит дегидрирование субстратов, образующихся в процессе метаболизма с переносом электронов и протонов на кофермент НАД (НАДФ).
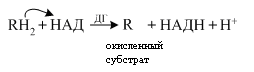
НАД - зависимая дегидрогеназа акцептор электронов и протонов от окисляемых веществ. Образующаяся восстановленная форма НАД затем окисляется при участии флавопротеидов, встроенных в мембрану митохондрий по схеме:.
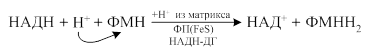
· В последующем электроны с восстановленной формы флавопротеидов переносятся при участии железосеросодержащих белков (Fe-S комплексов) на следующий компонент: КоQ по схеме:
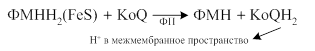
· КоQН2 окисляется системой цитохромов, на которые с КоQ перебрасываются только электроны, а протоны выталкиваются в межмембранное пространство:
· 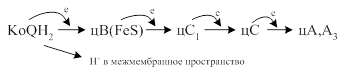
· Под действием цитохромоксидазы на молекулу кислорода перебрасываются 4 электрона с образованием восстановленной формы кислорода 2О2-, который в последующем взаимодействует с 4Н+ с образованием Н2О.
·
· 4.5.2. Короткая дыхательная цепь
· В короткой дыхательной цепи окисляются субстраты, для которых первичным акцептором электронов является флавопротеид (отсутствует этап окисления субстрата при участии НАД-ДГ). Веществами, окисляющимися в короткой цепи, являются, напимер, янтарная кислота, активные формы жирных кислот, глицерофосфат.
· Первая стадия окисления:
· 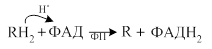
· В последующем ФАДН2 при участии комплекса флавопротеидов(FeS), окисляется КоQ:
· 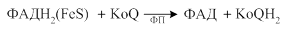
· Восстановленный КоQ далее окисляется также, как и в длиной дыхательной цепи, системой цитохромов.
·
· 4.5.3. Окислительные комплексы и их ингибиторы
· Длинная и короткая дыхательные цепи включают в свой состав структурно-функциональные фрагменты, которые называются окислительными (дыхательными) комплексами. В длинной цепи выделяют три основных комплекса (I, III, IV), а в короткой два (III, IV).
· 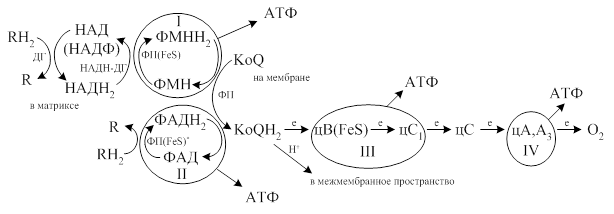
· I комплекс - НАДН - дегидрогеназный комплекс располагается между НАДН2 и КоQ и включает в себя ФП и FeS – белки.
· III комплекс - КоQН2-дегидрогеназный или (цитохром С - редуктазный комплекс) располагается между КоQ и цС и включает в себя цВ, FeS- белки, цС1.
· IV комплекс - цитохромоксидазный комплекс – окисляет цС и включает в себя цА,А3
· II дополнительный сукцинатдегидрогеназный комплекс включает ФП* и FeS*,
· Каждый дыхательный комплекс может быть выключен из работы дыхательной цепи определёнными веществами – ингибиторами, блокирующими передачу электронов и протонов.
· Ингибиторы первого комплекса – амитал, барбитураты, ротенон.
· Ингибитор второго комплекса – малонат.
· Ингибитор третьего комплекса – антимицин А.
Ингибиторы четвертого комплекса – Н2S, цианиды, СО.
Липиды пищи человека
Жиры. 70-100г в сутки. Содержат незаменимые полиненасыщенные жирные кислоты. Это линолевая кислота С18 :2, арахидоновая С20 :4. Они служат источником БАВ. Эйкозанойды. Полиненасыщенные содержатся в растительных маслах, поэтому в рационе четверть всех жиров должна быть представлена растительными жирами. У детей потребность в жирах выше чем у взрослых. При недостатке нарушается деятельность ЦНС, снижается иммунитет, нарушаются различные виды обмена, атеросклероз, ожирение.
Переваривание липидов
происходит в тонком кишечнике. Необходимыми условиями для этого являются:
- наличие ферментов;
- оптимальное рН;
- эмульгирование жиров.
. Эмульгирование повышает поверхность раздела липид / вода и обеспечивает большую поверхность контакта фермента и липида. В эмульгировании липидов основную роль играют жёлчные кислоты, выделяемые в просвет кишечника в составе жёлчи.
Различают простые и парные, первичные и вторичные жёлчные кислоты:
Простые жёлчные кислоты являются производными холановой кислоты.
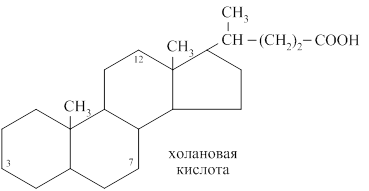
К простым жёлчным кислотам относятся холевая, дезоксихолевая кислота, хенодезоксихолевая и литохолевая кислоты.
Синтез жёлчных кислот из холестерина происходит в печени. Ключевым ферментом является 7 – α -гидроксилаза. Она переводит холестерин при участии цитохрома Р450 в 7-α - холестерин - 3,7 (ОН)2. Он, в свою очередь, переходит в хенодезоксихолевую кислоту - 3,7 (ОН)2 путём укорочения бокового радикала и в холевую кислоту - 3,7,12 (ОН)3. Эти две кислоты являются первичными жёлчными кислотами.
Полярность простых жёлчных кислот увеличивается при образовании парных жёлчных кислот путём присоединения глицина (гликокола) и таурина.
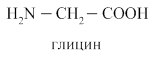
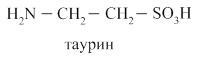
В кишечнике под действием микрофлоры происходит отцепление таурина, гликокола и ОН группы с образованием вторичных желчных кислот: дезоксихолевой и литохолевой.
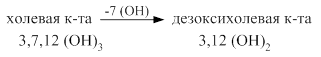
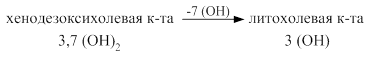
Все жёлчные кислоты имеют- Гидрофильне уч.-это ОН - группы, остатки таурина и гликокола, а гидрофобные – радикал жёлчной кислоты. Благодаря дифильности жёлчные кислоты располагаются в поверхностном слое жировой капли и уменьшают поверхностное натяжение.
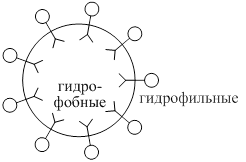
происходит дробление крупных капель липида на множество мелких – эмульгирование, резко возрастает поверхность соприкосновения липидных капель и ферментов.
Липолитические ферменты, участвующие в переваривании липидов, активны при pН 8 – 8,5. Основные ферменты переваривания липидов вырабатываются поджелудочной железой и стенкой тонкого кишечника.
Впереваривании ТАГ участвует поджелудочная липаза. Она вырабатывается в неактивной форме, и в тонком кишечнике взаимодействует с дополнительным белком колипазой, который повышает активность липазы и обеспечивает контакт фермента с соответствующими жирами. Поджелудочная липаза отщепляет последовательно остатки жирных кислот из α - положений с образованием β – моноацилглицерина (β -МАГ)
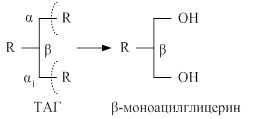
Образующиеся бета-МАГ в случае изомеризации в α –МАГ могут в дальнейшем подвергаться расщеплению под действием липазы до глицерина и жирных кислот. Около 50% МАГ подвергается всасыванию.
Переваривание глицерофосфолипидов происходит под действием ферментов фосфолипаз поджелудочной железы. Под действием фосфолипазы А2 отщепляется остаток жирной кислоты из β – положения с образованием продукта неполного распада глицерофосфолипида – лизофосфолипида, содержащего в β – положении гидрофильную гидроксильную группу. Лизофосфолипиды являются поверхностно активными веществами и усиливают процессы эмульгирования жиров. Лизофосфолипиды расщепляются лизофосфолипазой на жирную кислоту и глицерофосфохолин, который хорошо всасывается без гидролиза. Таким образом, при распаде глицерофосфолипидов образуются жирные кислоты и глицерофосфохолин.
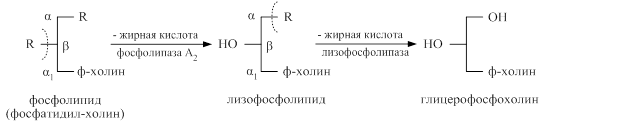
Эфиры холестерина расщепляются ферментом холестеролэстеразой.
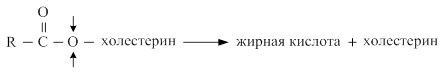
Переваривание сфинголипидов осуществляется ферментами эстеразами, фосфатазами, амидазами, гликозидазами.