




Закон был открыт Н.Е. Введенским, который, изучая процессы возбуждения на нервно-мышечном препарате лягушки, показал, что после развития утомления в синапсе нерв еще долго может генерировать нервные импульсы в ответ на раздражение рецепторов, что связано с очень низким обменом веществ в нерве и большой интенсивностью процессов ресинтеза.
6 вопрос Структурно-функциональные особенности возбуждающих и тормозящих синапсов и механизм проведения возбуждения через них. Каковы особенности постсинаптической мембраны. Понятие о медиаторе, его роли в проведении возбуждения через синапс, виды медиаторов. Различия в действии возбуждающих и тормозных медиаторов. Роль холинстеразы в работе синапсов. Что такое впсп и тпсп и каковы их особенности.
Взаимодействие нейронов между собой (и с эффекторными органами) происходит через специальные образования — синапсы (греч. — контакт). Они образуются концевыми разветвлениями нейрона на теле или отростках другого нейрона. Чем больше синапсов на нервной клетке, тем больше она воспринимает различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия в разнообразных реакциях организма. Особенно много синапсов в высших отделах нервной системы и именно у нейронов с наиболее сложными функциями.
В структуре синапса различают три элемента (рис. 1):
- пресинаптическую мембрану, образованную утолщением мембраны конечной веточки аксона;
- синаптическую щель между нейронами;
- постсинаптическую мембрану — утолщение прилегающей поверхности следующего нейрона.
|
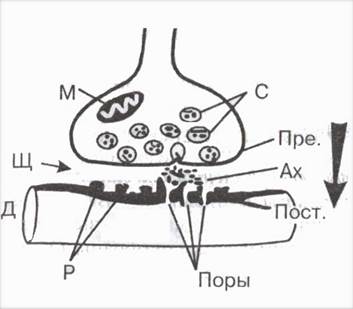
В большинстве случаев передача влияния одного нейрона на другой осуществляется химическим путем. В пресинаптической части контакта имеются синаптические пузырьки, которые содержат специальные вещества — медиаторы или посредники. Ими могут быть ацетилхолин (в некоторых клетках спинного мозга, в вегетативных узлах), норадреналин (в окончаниях симпатических нервных волокон, в гипоталамусе), некоторые аминокислоты и др. Приходящие в окончания аксона нервные импульсы вызывают опорожнение синаптических пузырьков и выведение медиатора в синаптическую щель.
По характеру воздействия на последующую нервную клетку различают возбуждающие и тормозящие синапсы.
В возбуждающих синапсах медиаторы (например, ацетилхолин) связываются со специфическими макромолекулами постсинаптической мембраны и вызывают ее деполяризацию. При этом регистрируется небольшое и кратковременное (около 1 мс) колебание мембранного потенциала в сторону деполяризации или возбуждающий постсинаптический потенциал (ВПСП). Для возбуждения нейрона необходимо, чтобы ВПСП достиг порогового уровня. Для этого величина деполяризационного сдвига мембранного потенциала должна составлять не менее 10 мВ. Действие медиатора очень кратковременно (1 -2 мс), после чего он расщепляется на неэффективные компоненты (например, ацетилхолин расщепляется ферментом холинэстеразой на холин и уксусную кислоту) ил и поглощается обратно пресинаптическими окончаниями (например, норадреналин).
В тормозящих синапсах содержатся тормозные медиаторы (например, гамма-аминомасляная кислота). Их действие на постсинаптическую мембрану вызывает усиление выхода ионов калия из клетки и увеличение поляризации мембраны. При этом регистрируется кратковременное колебание мембранного потенциала в сторону гиперполяризации — тормозящий постсинаптический потенциал (ТПСП). В результате нервная клетка оказывается заторможенной. Возбудить ее труднее, чем в исходном состоянии. Для этого понадобится более сильное раздражение, чтобы достичь критического уровня деполяризации.
7.Понятие о нервных центрах
Нервный центр — совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт. Представление о структурно-функциональной основе нервного центра обусловлено историей развития учения о локализации функций в центральной нервной системе. На смену старым теориям об узкой локализации, или эквипотенциальности, высших отделов головного мозга, в частности коры большого мозга, пришло современное представление о динамической локализации функций, основанное на признании существования четко локализованных ядерных структур нервных центров и менее определенных рассеянных элементов анализаторных систем мозга. При этом с цефализацией нервной системы растут удельный вес и значимость рассеянных элементов нервного центра, внося существенные различия в анатомических и физиологических границах нервного центра. В результате функциональный нервный центр может быть локализован в разных анатомических структурах. Например, дыхательный центр представлен нервными клетками, расположенными в спинном, продолговатом, промежуточном мозге, в коре большого мозга.
Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований.
1. Односторонность проведения возбуждения. В рефлекторной дуге, включающей нервные центры,
процесс возбуждения распространяется в одном направлении (от входа, афферентных путей к выходу, эфферентным путям).
2. Иррадиация возбуждения. Особенности структурной организации центральных нейронов, огромное
число межнейронных соединений в нервных центрах существенно модифицируют (изменяют) направление распространения процесса возбуждения в зависимости от силы раздражителя и функционального состояния центральных нейронов. Значительное увеличение силы раздражителя приводит к расширению области вовлекаемых в процесс возбуждения центральных нейронов — иррадиации возбуждения.
3. Суммация возбуждения. В работе нервных центров значительное место занимают процессы пространственной и временной суммации возбуждения, основным нервным субстратом которой является постсинаптическая мембрана. Процесс пространственной суммации афферентных потоков возбуждения облегчается наличием на мембране нервной клетки сотен и тысяч синаптических контактов. Процессы временной суммации обусловлены суммацией ВПСП на постсинаптической мембране.
4. Наличие синаптической задержки. Время рефлекторной реакции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс. При относительно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка). В нервных клетках высших животных и человека одна синаптическая задержка примерно равна 1 мс. Если учесть, что в реальных рефлекторных дугах
имеются десятки последовательных синаптических контактов, становится понятной длительность большинства рефлекторных реакций — десятки миллисекунд.
Высокая утомляемость. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением. Этот процесс связан с деятельностью синапсов — в последних наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, происходит адаптация постсинаптического рецептора к медиатору.
6. Тонус. Тонус, или наличие определенной фоновой активности нервного центра, определяется тем, что в покое в отсутствие специальных внешних раздражений определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки. Даже во сне в высших отделах мозга остается некоторое количество фоновоактивных нервных клеток, формирующих «сторожевые пункты» и определяющих некоторый тонус соответствующего нервного центра.
7. Пластичность. Функциональная возможность нервного центра существенно модифицировать картину осуществляемых рефлекторных реакций. Поэтому пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами.
8. Конвергенция. Нервные центры высших отделов мозга являются мощными коллекторами, собирающими разнородную афферентную информацию. Количественное соотношение периферических рецепторных и промежуточных центральных нейронов (10:1) предполагает значительную конвергенцию («сходимость») разномодальных сенсорных посылок на одни и те же центральные нейроны. На это указывают прямые исследования центральных нейронов: в нервном центре имеется значительное количество поливалентных, полисенсорных нервных клеток, реагирующих на разномодальные стимулы (свет, звук, механические раздражения и т. д.). Конвергенция на клетках нервного центра разных афферентных входов предопределяет важные интегративные, перерабатывающие информацию функции центральных нейронов, т. е. высокий уровень интеграционных функций. Конвергенция нервных сигналов на уровне эфферентного звена рефлекторной дуги определяет физиологический механизм принципа «общего конечного пути» по Ч. Шеррингтону.
9. Интеграция в нервных центрах. Важные интегративные функции клеток нервных центров ассоциируются с интегративными процессами на системном уровне в плане образования функциональных объединений отдельных нервных центров в целях осуществления сложных координированных приспособительных целостных реакций организма (сложные адаптивные поведенческие акты).
10. Свойство доминанты. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в центральной нервной системе. По А.А.Ухтомскому, доминантный нервный очаг характеризуется такими свойствами, как повышенная возбудимость, стойкость и инертность возбуждения, способность к суммированию возбуждения.
В доминантном очаге устанавливается определенный уровень стационарного возбуждения, способствующий суммированию ранее подпороговых возбуждений и переводу на оптимальный для данныхусловий ритм работы, когда этот очаг становится наиболее чувствительным. Доминирующее значение такого очага (нервного центра) определяет его угнетающее влияние на другие соседние очаги возбуждения. Доминантный очаг возбуждения «притягивает» к себе возбуждение других возбужденных зон (нервных центров). Принцип доминанты определяет формирование главенствующего (активирующего) возбужденного нервного центра в тесном соответствии с ведущими мотивами, потребностями организма в конкретный момент времени.
11. Цефализация нервной системы. Основная тенденция в эволюционном развитии нервной системы проявляется в перемещении, сосредоточении функции регуляции и координации деятельности организма в головных отделах ЦНС. Этот процесс называется цефализацией управляющей функции нервной системы. При всей сложности складывающихся отношений между старыми, древними и эволюционно новыми нервными образованиями стволовой части мозга общая схема взаимных влияний может быть представлена следующим образом: восходящие влияния (от нижележащих «старых» нервных структур к вышележащим «новым» образованиям) преимущественно носят возбуждающий стимулирующий характер, нисходящие (от вышележащих «новых» нервных образований к нижележащим «старым» нервным структурам) носят угнетающий тормозной характер. Эта схема согласуется с представлением о росте в процессе эволюции роли и значения тормозных процессов в осуществлении сложных интегративных рефлекторных реакций.
В основу классификации положены морфологический и функциональный критерий. По локализации в структурах нервной системы различают корковые, подкорковые и спинальные центры. В головном мозге также выделяют центры диэнцефальные, мезэнцефальные, бульбарные, гипоталамические, таламические.
На функциональной основе центры нервной системы разделяют по регулируемой функции (например, сосудодвигательный центр, центр теплообразования, дыхательный центр и др.) или по афферентному восприятию (например, центры зрения, слуха, обоняния и др.). Выделяют также центры нервной системы, которые формируют мотивационные состояния организма, являясь пейсмекерами мотивационных возбуждений (центры голода, жажды, насыщения и др.). Существуют центры нервной системы, которые на основе интеграции возбуждений формируют целостные реакции организма (центры глотания, чихания, дефекации, половой центр и др.).
Интегративная деятельность спинного мозга связана со структурной организацией его сегментов. В пределах каждого сегмента спинного мозга возможна интеграция простейших двигательных реакций.
Миотатический рефлекс - одна из простых реакций на растяжение мышцы, осуществляемая на уровне спинного мозга. Реакция растяжения является основой регуляции длины мышцы, изменение которой возникает при поднятии груза или при сокращении мышцы антагониста.
Сухожильные рефлексы в противоположность реакции на растяжение мышцы способствуют быстрому ее расслаблению. Они защищают ее от повреждения при сильных сокращениях и участвуют в регуляции напряжения в мышце.
Стабилизируя подвижные звенья скелета, миотатические и сухожильные рефлексы создают основу для осуществления непроизвольных и произвольных движений.
Защитные рефлексы спинного мозга возникают при раздражении кожной поверхности повреждающими раздражителями. В этом случае при раздражении рецепторов кожи происходит отдергивание конечности, или сгибательный рефлекс.
Спинальные двигательные центры находятся под контролирующим влиянием вышележащих отделов центральной нервной системы.

